Downloaded 697 times






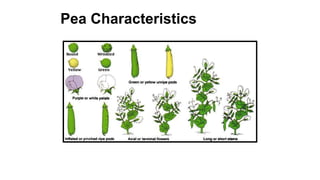



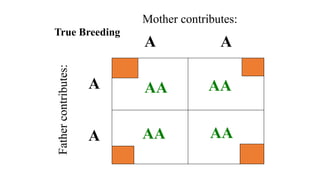
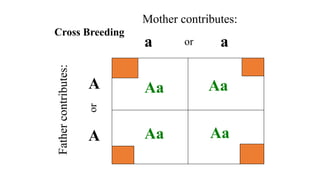
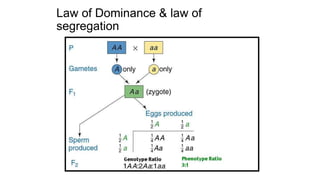
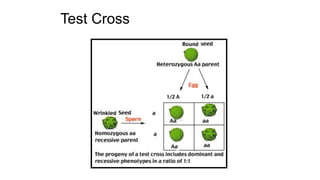







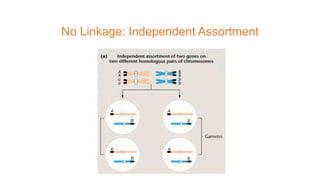

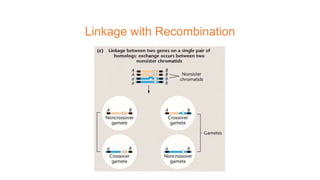
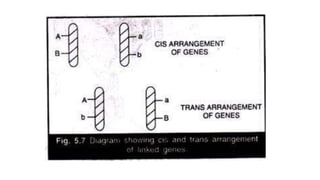

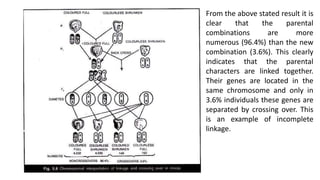


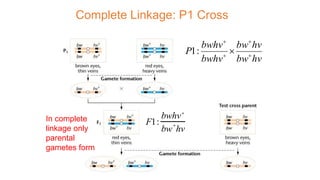
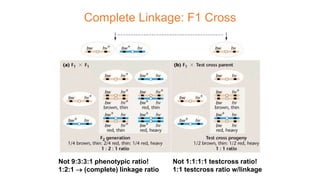





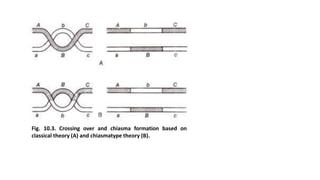


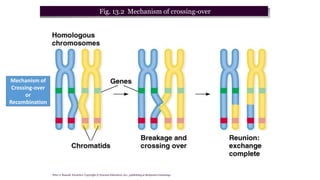



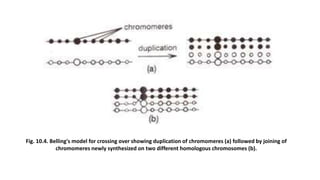




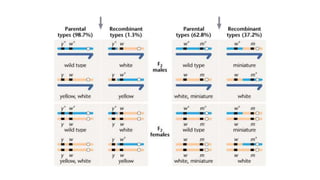
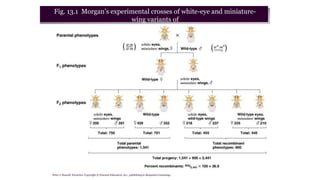




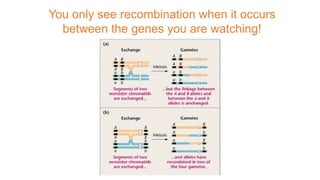
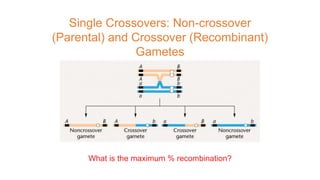
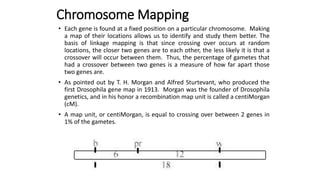



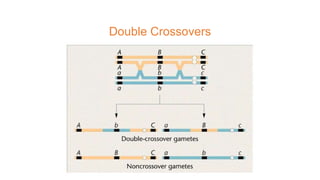


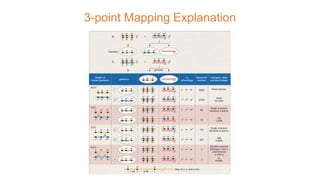







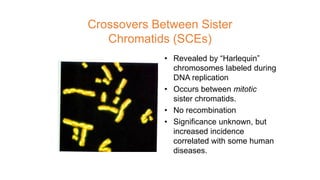

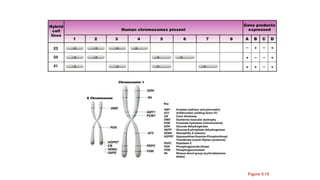


The document discusses the principles of genetic inheritance as founded by Gregor Mendel, including key concepts such as genes, alleles, and phenotypes, alongside Mendel's hypotheses on dominance, segregation, and independent assortment. It also delves into the ideas of genetic linkage, complete and incomplete linkage, and the significance of crossing over, referring to various experiments and observations by Mendel, Bateson, Punnet, and Morgan. Finally, it explains the mechanisms of crossing over and theories surrounding genetic recombination, emphasizing their role in inheritance.
























































![Krishna Cv[1]](https://cdn.slidesharecdn.com/ss_thumbnails/krishnacv1-12777037042611-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)



















