Downloaded 147 times



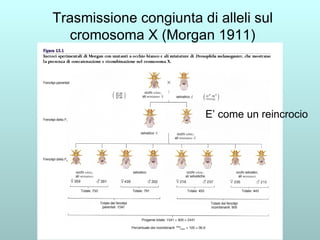

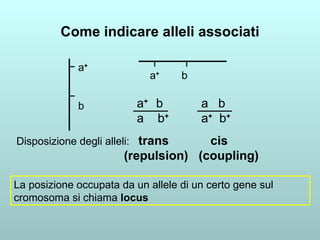
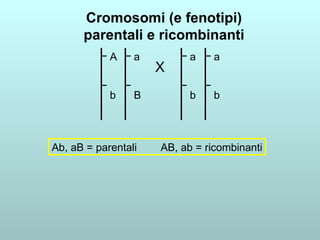
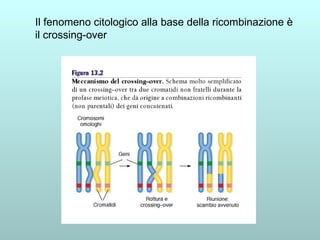

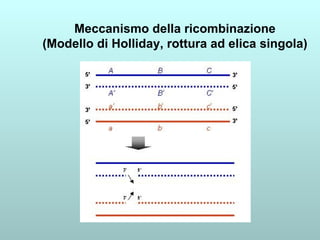
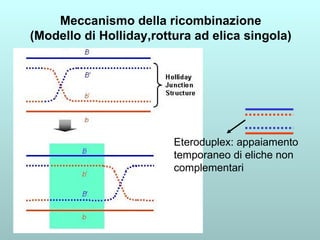
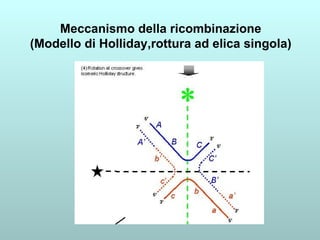
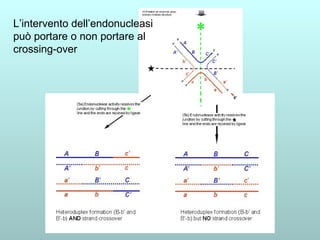
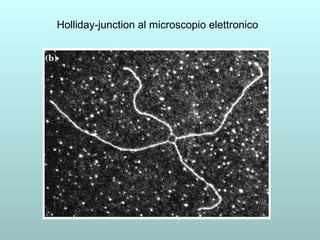
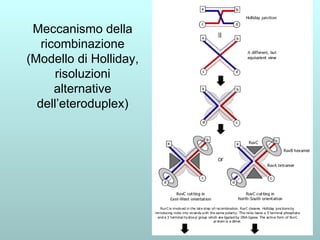
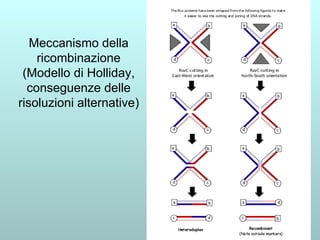
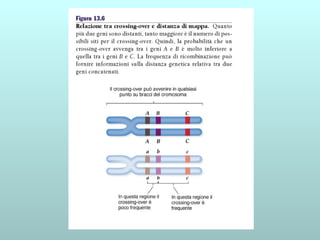
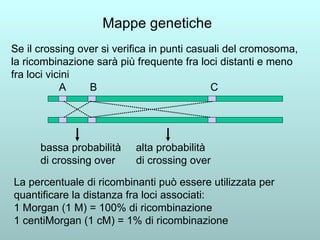



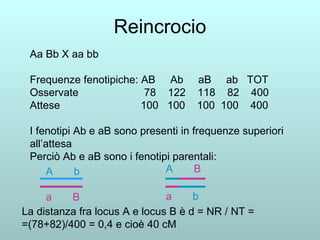

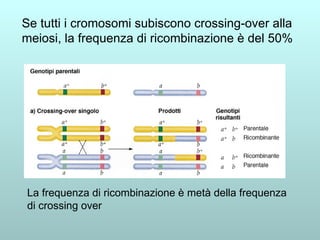








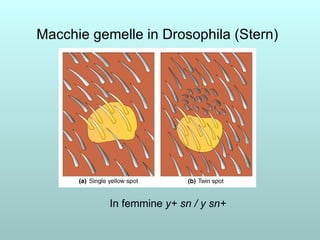

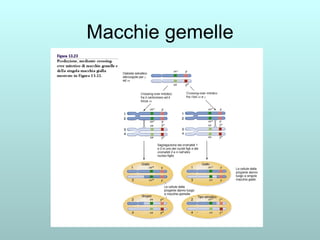

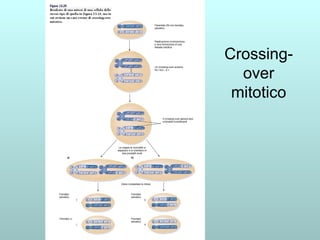
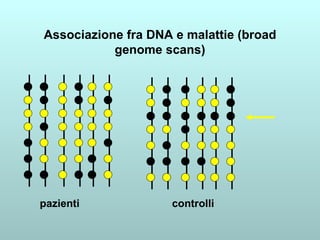
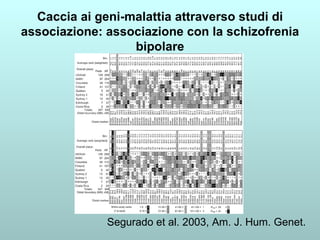

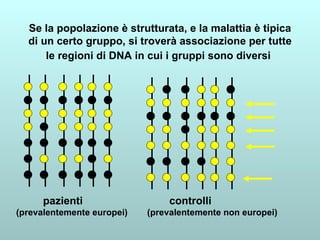
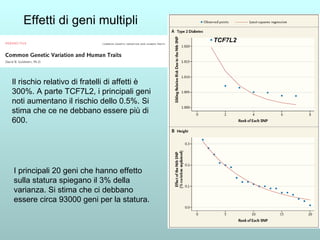






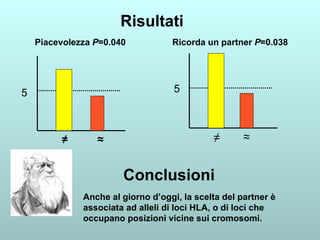
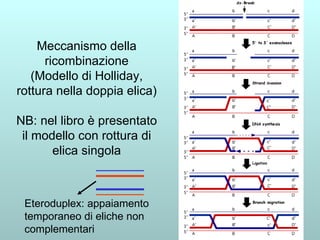

Il capitolo 13 discute la mappatura dei geni negli eucarioti, evidenziando che gli alleli non sempre vengono trasmessi in modo indipendente a causa dell'associazione dei loci sui cromosomi. Vengono descritti i meccanismi di ricombinazione, come il crossing-over, e si spiega come calcolare la distanza tra loci basandosi sulla frequenza di ricombinazione. Infine, viene trattata l'associazione tra varianti genetiche e malattie, illustrando come l'eterogeneità genetica possa influenzare i risultati degli studi di associazione.




























































