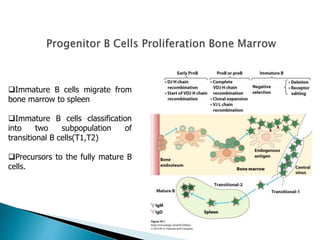
B cells develop from hematopoietic stem cells in the bone marrow over 1-2 weeks. They undergo gene rearrangement and selection to produce a functional B cell receptor. Immature B cells migrate to the spleen where they further differentiate into transitional and then mature B cells. Upon antigen activation, B cells proliferate and differentiate into plasma cells that secrete antibody or memory B cells. The processes of somatic hypermutation and class switching increase antibody affinity and modify isotype.