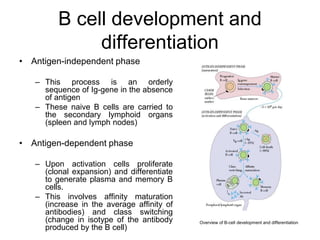
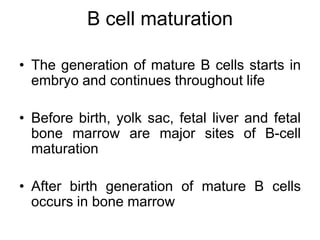
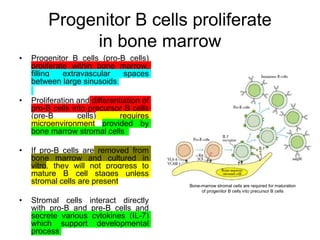
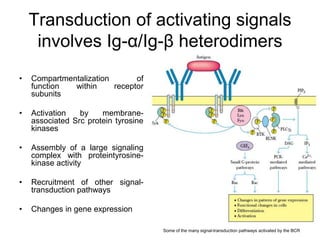
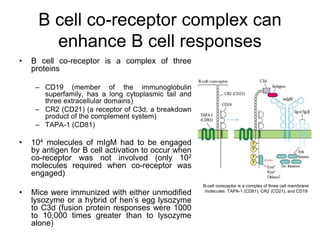
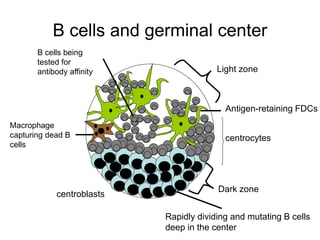
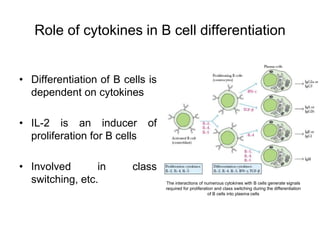
The document summarizes B cell development, activation, and differentiation. It is divided into three main stages: 1) generation of mature B cells, 2) activation of mature B cells, and 3) differentiation into plasma and memory B cells. The process involves an antigen-independent maturation phase and an antigen-dependent activation and differentiation phase where B cells proliferate, undergo affinity maturation and class switching in germinal centers. Cytokines also play a role in B cell differentiation.




































































![Polymer [ बहुलक ] Chemistry Notes PDF - Irfanullah Mehar - JJ Sir Chemistry.pdf](https://cdn.slidesharecdn.com/ss_thumbnails/polymerchemistrynotespdf-irfanullahmehar-jjsirchemistry-260210172118-3f9b37f7-thumbnail.jpg?width=640&height=640&fit=bounds)







