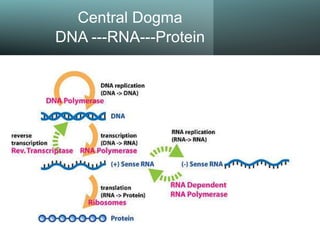
3. Cấu trúc DNA
Các khối cấu tạo của DNA là các nucleotide. Mỗi nucleotide được tạo thành từ một đường,
một nhóm phốt phát và một bazơ nitơ. Đường là deoxyribose trong DNA và ribose trong
RNA.
4. Double Helix, Liên kết Phosphodiester,
và các rãnh chính và phụ
DNA có (a) cấu trúc xoắn kép và (b) liên kết phosphodiester. (C)
các rãnh chính và rãnh phụ là các vị trí liên kết với các protein liên
kết DNA trong các quá trình như phiên mã (sao chép RNA từ
DNA) và sao chép.
5. Thông tin Di truyền
Trình tự của bốn base cung cấp
tất cả thông tin cần thiết để xây
dựng bất kỳ cơ thể sống nào.
Các base bổ sung tạo thành liên
kết hydro với nhau trong chuỗi
xoắn kép.
Các base lớn hơn (purin) bắt
cặp với các base nhỏ hơn
(pyrimidin).
Điều này giữ cho chiều rộng của
chuỗi xoắn kép không đổi. Cụ
thể hơn, A ghép với T và C ghép
với G.
7. DNA phát triển ở 15N (dải màu xanh) nặng hơn DNA được phát triển ở 14N (dải màu
đỏ), và lắng xuống mức thấp hơn trong quá trình siêu ly tâm. Sau một vòng sao chép,
DNA lắng đọng một nửa giữa mức 15N và 14N (dải màu tím), loại trừ mô hình sao chép
bảo thủ. Sau lần nhân rộng thứ hai, mô hình nhân rộng phân tán đã bị loại trừ. Những
dữ liệu này đã hỗ trợ mô hình sao chép bán bảo tồn.
8. Các đặc tính của sao chép DNA
Mostly in E. coli, but many features are general
1. Sự sao chép xảy ra theo hai hướng
2. Sợi đôi của phân tử DNA phải được tháo xoắn
3. Sao chép theo kiểu nửa gián đoạn ( sợi liên tục
và sợi không liên tục)
Leading strand is formed continuously
Lagging strand is formed from Okazaki fragments - discovered by
Tuneko and Reiji Okazaki
9. Figure 28.5
Bidirectional
replication of the E.
coli chromosome.
(a) If replication is
bidirectional, auto-
radiograms of
radioactively
labeled replicating
chromosomes
should show two
replication forks
heavily labeled with
radioactive
thymidine. (b) An
autoradiogram of
the chromosome
from a dividing E.
coli cell shows
bidirectional
replication. (Photo
courtesy of David
M. Prescott,
University of
Colorado.)
12. Tại điểm khởi đầu sao chép,
topoisomerase II làm giãn nhiễm
sắc thể siêu cuộn. Hai nhánh sao
chép được hình thành do sự mở
của DNA sợi kép tại ori, và helicase
tách các sợi DNA, được bao bọc
bởi các protein liên kết sợi đơn để
giữ cho các sợi được tách ra. Quá
trình nhân đôi DNA xảy ra theo cả
hai hướng. Một đoạn mồi RNA bổ
sung cho sợi cha mẹ được tổng
hợp bởi RNA primase và được kéo
dài bởi DNA polymerase III thông
qua việc bổ sung nucleotide vào
đầu 3'-OH. Trên sợi dẫn đầu, DNA
được tổng hợp liên tục, trong khi
trên sợi lagging, DNA được tổng
hợp thành các đoạn ngắn được gọi
là đoạn Okazaki. Các đoạn mồi
RNA trong sợi lagging bị loại bỏ bởi
hoạt động exonuclease của DNA
polymerase I, và các đoạn Okazaki
được nối với DNA ligase.
13. Tổng hợp
DNA
1.Tổng hợp bắt đầu từ “khởi điểm-
origin”.
• Khởi điểm ở E.coli là một đoạn
240-300 cặp nucleotid (baz), ở vi
rút SV40 ký sinh trên động vật có
khởi điểm dài 65 cặp nucleotid.
• Tế bào chân hạch có nhiều khởi
điểm gọi là “trình tự nhân đôi độc
lập”
2. Cơ chế tổng hợp
1). Các enzim tham gia
2). Cơ chế
14. 14
KHỞI ĐIỂM SAO CHÉP DNA “ORI”
Sinh vật sơ hạch có một
khởi điểm sao chép
Sinh vật chân hạch có nhiều
khởi điểm sao chép
15. 2.Cơ chế tổng hợp
1). Các enzim tham gia Nhiệm vụ
- dnaA,dnaB(helicaz) và dnaC Tháo vòng xoắn tại khởi điểm
- Primaz (RNA polymerase) Tổng hợp ARN prime
- Helicaze Tháo xoắn
- SSB Giữ các polynucleotid duỗi ra
- DNA gyraz (Topoisomerase II) Xoắn DNA theo chiều xoắn âm
- DNA polymeraz III Tổng hợp các phần chính của
DNA
-DNA polymeraz I Loại bỏ ARN prime và nối các
đoạn Okazaki
- DNA ligaz Nối các đầu DNA lại
16. (a) Mô hình sách giáo khoa chuẩn về một tổ hợp DNA cho thấy các quá trình kết hợp và phối hợp
cao của quá trình tổng hợp sợi leading và sợi lagging. DNA polymerase III được kết nối với DnaB
helicase thông qua tiểu đơn vị của bộ nạp-kẹp và hai hoặc ba lõi polymerase sao chép đồng thời
DNA từ cả mẫu DNA sợi leading và lagging. SsDNA trong vòng lặp sợi trễ được liên kết bởi các
protein liên kết ssDNA (SSB). (b) Các nghiên cứu gần đây đã chỉ ra rằng DNA polymerase III của
E. coli có thể dễ dàng trao đổi ở ngã ba và quá trình tổng hợp sợi leading và lagging có thể không
được kết hợp chặt chẽ, hoặc thậm chí có thể được thực hiện bởi các holoenzyme DNA
polymerase III khác nhau. DnaB helicase cũng có thể được tách khỏi phức hợp DNA polymerase
và chuyển vị trí trước đỉnh của ngã ba.
17.
18. Cơ chế tổng hợp
Các enzime tham gia:
Năm 1957, Arthur Kornberg chứng minh có sự
tồn tại của - DNA polymerase I
DNA Polymerase I có 3 vùng hoạt động khác
nhau nằm trong cùng một sợi polypeptid:
N- 5’ 3’ Exonucleaz 3’ 5’ Exonuleaz Polymeraz -C
1/ 5’ 3’
Polymera
se: Hoạt
động tổng
hợp đoạn
ngắn DNA
2/ 3’
5’
exonuclea
z: cắt bỏ
nucleotid
ở đầu 3’
3/ 5’ 3’
exonuclea
z: cắt
nucleotid
ở đầu 5
19. Tại sao cần có hoạt động của 3’ 5’ exonucleaz ?
• 3’ 5’ exonucleaz phục vụ chức năng đọc và
sửa sai
• Nó loại bỏ những baz bắt cặp sai để cho DNA
polymerase tổng hợp lại.
20. Hoạt động đọc và sửa sai của 3’
5’ exonucleaz
• DNAPI dừng nếu Nu sai được
thêm vào - nó không thể thêm Nu
tiếp theo trong chuỗi
• Hoạt động sửa lỗi diễn ra chậm
so với tổng hợp, nhưng sự
ngưng trệ của DNA PI sau khi
chèn một Nu không chính xác
cho phép hoạt động sửa lỗi bắt
kịp với quá tổng hợp và loại bỏ
Nu không chính xác.
22. Tại sao cần có hoạt động của 5’ 3’ exonucleaz?
• Hoạt động của 5’ 3’ exonucleaz được sử dụng để cắt bỏ
RNA primers trong phản ứng gọi là “nick translation”
• DNA Pol I còn có chức năng năng sửa sai tại vùng 5’-3’
23. DNA Polymeraz I 5’ 3’exonucleaz
DNA Pol I còn có chức năng năng sửa sai tại vùng 5’-3’
24. DNA
Polymerase I
có phải là
enzyme sao
chép chính
không ?
Năm 1969, John Cairns và Paula
deLucia đã phân lập được một dòng vi
khuẩn đột biến chỉ có 1% hoạt tính
DNAP I (polA)
Vi khuẩn đột biến siêu nhạy cảm với bức
xạ UV
Nhưng đột biến vẫn ổn, tức là nó có thể
phân chia, vì vậy rõ ràng là nó có thể
nhân đôi DNA của mình
Sự kết luận: DNAP I KHÔNG phải là
enzym sao chép chính ở E. coli
25. Các manh
mối
khác….
- DNAP I quá chậm (600 dNTP được thêm vào / phút -
sẽ mất 100 giờ để tái tạo bộ gen thay vì 40 phút)
- DNAP I chỉ xử lý vừa phải
(Tiến trình đề cập đến số lượng dNTP được thêm vào
chuỗi DNA đang phát triển trước khi enzyme phân ly
khỏi khuôn DNA mẫu)
Kết luận:
Phải có DNA polymerase bổ sung.
Các nhà hóa sinh đã phân lập chúng từ chủng đột biến
polA
26. DNA
Polymerase
III
Polymerase sao chép "thực" ở E. coli
Nhanh chóng: lên đến 1.000 dNTP
được thêm vào / giây / enzyme
Nó có tính xử lý cao:> 500.000 dNTP
được thêm vào trước khi phân tách
Chính xác: tạo ra lỗi 1/107 dNTP
được thêm vào, với tính năng sửa lỗi,
tỷ lệ lỗi cuối cùng nói chung là 1/1010.
27. -Thành phần: Ở E.coli có kích
thước 800 kdalton và gồm 8
sợi polypeptit (alpha),
(epsilon), (theta), (beta),
(tau), (gamma), (delta), ’
- Chức năng: Tổng hợp phần
chính của DNA: sợi
polynucleotid liên tục mới và
các đoạn Okazaki
• : xúc tác tổng hợp sợi không
liên tục (Okazaki); : xúc tác
tổng hợp các đoạn liên tục; :
có hoạt tính 3’ 5’
exonucleaz; : gắn lõi làm bởi
, và vào sợi polynucleotid
làm khuôn
DNA Polymeraz III
29. Cơ chế sao chép DNA ở E.coli
Gồm 3 giai đoạn
• Khởi đầu
• Kéo dài
• Kết thúc
Hướng sao chép: 5’ 3’
30. 1. Tổng hợp bắt đầu từ một khởi điểm duy nhất (240-300
cặp nucleotid và tiến hành theo hai chiều ngược nhau).
2. Khởi động bộ máy:
+Protein dnaA bám vào bốn vùng đặc biệt (mỗi vùng chứa 9
cặp nucleotid) của khởi điểm
+ Phức hợp protein dnaB và dnaC kết với dnaA mở xoắn
DNA, dnaC có lẽ có chức năng tải dnaB đến khởi điểm;
dnaB có hoạt tính của helicaz tháo rời hai sợi
polynucleotid để polymeraz bám vào
+ Một nhóm SSB protein giữ cho các sợi polynucleotid, vừa
mới bị tách rời, duỗi thẳng ra
Khởi đầu
31. Arrangement of sequences in the E. coli replication origin, oriC
The E. coli replication origin is known as oriC. In E. coli, the oriC
consists of 13 mer repeats followed by 9 mer repeats.
A protein, DnaA would bind to the 9 mer repeats, and the DNA would
then coil around the protein complex (many DnaA) forming a protein
core.
This coiling stimulates the AT rich region in the 13 mer sequence to
unwind, thus allowing enzymes and other factors to bind and
replication would start.
32. Mô hình khởi động tổng hợp tại khởi điểm (ở E.coli)
Ba đoạn chứa13 cặp baz: 5’-GATCTNTTNTTTT-3’
3’-CTAGANAANAAAA-5’
Vùng giàu AT: dễ tháo xoắn
33. Mô hình khởi động tổng hợp tại khởi điểm (ở E.coli)
34. Cả hai sợi polynucleotid làm khuôn tổng hợp hai sợi
polynucleotid mới. Để tiến hành tổng hợp, enzim primaz
dùng các sợi polynucleotid cũ làm khuôn tổng hợp một
đoạn ARN dài 6-15 nucleotid gọi là đầu mồi-prime
(primer). DNA polymeraz gắn nucleotid đầu tiên vào đầu
3’ của prime
Khởi đầu (tt)
Tộng hợp DNA cần
một đoạn mồi
(prime) ARN để khởi
đầu
36. 36
Vai trò của đoạn mồi?
Cung cấp 3’-OH giúp DNA polymerase kéo dài sợi mới
37. Tổng hợp xong một mồi, đó là lúc sự kéo dài chuỗi DNA
mới bắt đầu. Ở E. coli, enzyme polymerase III thực hiện
chức năng then chốt này. Tốc độ tổng hợp trung bình là
1.000 nucleotide mỗi giây.
Trên sợi khuôn dẫn đầu (3'→5'): Trước tiên, một đoạn mồi
RNA được tổng hợp với đầu 3'-OH tự do. Sau đó, enzyme
DNA polymerase III (replisome) bắt đầu kéo dài chuỗi
DNA mới sinh trưởng theo chiều 5'→3' một cách liên tục.
Kéo dài
http://www.learnerstv.com/animation/biology/replication.swf
38. Nếu DNA polymerase chỉ tổng hợp từ 5 ’đến 3’
thì các ngã ba sao chép di chuyển có hướng
như thế nào?
39.
40. Trên sợi khuôn ra chậm (5'→3'): Sự kéo dài diễn ra không liên tục dưới
dạng các đoạn Okazaki. Kích thước trung bình mỗi đoạn Okazaki ở E.
coli là 1.000 - 2.000 nucleotide; ở eukaryote là 100-200
Quá trình này đòi hỏi sự "mồi hóa" nhiều lần và có tính chu kỳ, với sự
tham gia lần lượt của bốn enzyme như sau:
• Primase tổng hợp một đoạn mồi RNA
• DNA polymerase III hoàn chỉnh kéo dài đoạn Okazaki
• DNA polymerase I vừa cắt bỏ dần từng nucleotide của đoạn mồi vừa
lấp khoảng trống bằng cách kéo dài dần đoạn Okazaki theo sau
• DNA ligase hàn liền khe hở còn lại giữa hai đoạn Okazaki kề nhau
bằng một liên kết 3',5'-phosphodieste
Kéo dài
41. - Cắt bỏ RNA mồi đầu tiên của sợi liên tục
- Để hoàn thành việc tái bản DNA, tế bào phải lấp đầy các
khoảng trống do các RNA mồi bị cắt bỏ để lại
-Termination: the "ter" locus, rich in Gs and Ts, signals the end of replication. A
Ter protein is also involved. Ter protein is a contrahelicase and prevents
unwinding
Kết thúc
Xem animation DNA replication 1
42.
43.
44. Tại sao đoạn mồi phải loại bỏ sau khi DNA sao chép xong?
45. Trên RNA primer trình tự U bắt cặp được với A trên DNA ,
đầu 5’ có 3 gốc Phosphat
A = U
46. Hãy cho biết chức năng của các enzyme tham gia sao chép DNA?
48. Tổng hợp DNA ở tế bào chân hạch
Một tế bào di chuyển qua một
loạt các giai đoạn một cách có
trật tự. Trong giai đoạn
interphase, G1 liên quan đến sự
phát triển tế bào và tổng hợp
protein, giai đoạn S liên quan đến
quá trình sao chép DNA và sao
chép của trung thể, và G2 liên
quan đến sự phát triển thêm và
tổng hợp protein. Giai đoạn
nguyên phân diễn ra sau giữa
các pha. Nguyên phân là sự phân
chia nhân trong đó các nhiễm sắc
thể đã nhân đôi phân ly và phân
bố thành các nhân con. Thông
thường tế bào sẽ phân chia sau
khi nguyên phân trong một quá
trình gọi là cytokinesis, trong đó
tế bào chất được phân chia và
hai tế bào con được hình thành.
49. 1.Chu kỳ phân cắt đẳng nhiễm
+ Giai đoạn nghỉ G1( 30-40% thời gian giữa hai lần
phân
cắt): tổng hợp protein và ARN làm tế bào to thêm
+ Giai đoạn S (30-50% thời gian chu kỳ phân
cắt):Tổng hơp DNA và protein histon, chia nhiễm sắc
thể ra làm hai bộ tương đồng
+ Giai đoạn G2 (5-20% chu kỳ phân cắt): Chuẩn
bị phân cắt
+ Giai đoạn M và D: Tiến hành phân cắt cho ra
hai tế bào con
Tổng hợp DNA ở tế bào chân hạch
50. Hàng ngàn điểm sao chép cùng lúc; điểm sao chép điển
hình: 200,000 ~ 300,000 bp in length
Sao chép trên nhiễm sắc thể Eukaryote
51. Chu kỳ tăng trưởng và phân cắt của tế bào
G1:- Giai đoạn nghỉ
chiếm 30-40% thời
gian chu kỳ phân cắt.
G2:
Giai đoạn G2 chiếm 5-
10% thời gian chu kỳ
sinh trưởng.
Ty thể và các cấu trúc
khác được nhân đôi
Một số protein cũng
được tổng hợp
M: Phân nhân
(Tiền,Biến,Tiến và
Chung kỳ)
52. Origins of Replication in Eukaryotes
Tổ chức điểm khởi đầu, đặc điểm và sự kích hoạt
ở sinh vật nhân chuẩn phức tạp hơn so với giới vi
khuẩn hoặc sinh vật cổ và sai lệch đáng kể so với
mô hình được thiết lập để bắt đầu sao chép nhân
sơ.
54. Tổng hợp DNA ở tế bào chân hạch
(1) Protein T bám vào khởi điểm và cùng với “yếu
tố nhân đôi-RFA” mở xoắn DNA tại khởi điểm
(2) Phản ứng tổng hợp tiến hành theo hai chiều
ngược nhau, khởi đầu từ một ARN prime, do
primaz xúc tác. Một loại polymeraz III ( Pol)
xúc tác tổng hợp đoạn polynucleotid liên tục
trong khi pol phối hợp với primaz tổng hợp
sợi không liên tục (Okazaki).
55. Phức hợp sao chép DNA ở sinh vật nhân chuẩn. Sao chép trên sợi leading và sợi
lagging được thực hiện lần lượt bởi Pol ε và Pol δ. Nhiều yếu tố tái tạo (bao gồm
FPC [phức hợp bảo vệ ngã ba], Claspin, And1 và RFC [yếu tố sao chép C kẹp tải])
chịu trách nhiệm điều chỉnh các chức năng polymerase và phối hợp tổng hợp DNA
với việc tháo sợi khuôn bằng Cdc45-MCM [mini -bảo trì nhiễm sắc thể] -GINS [go-
ichi-ni-san]. Hệ thống sao chép cũng liên kết với các protein điểm kiểm tra như các
cơ chế sao chép DNA và giám sát tính toàn vẹn của bộ gen.
56. Mô hình cơ chế tổng
hợp ở vi rút SV40 (Simian
virus 40)
63. The telomeres of humans consist of as many as 2000 repeats of the sequence
5' TTAGGG 3'.
5'...TTAGGG TTAGGG TTAGGG TTAGGG TTAGGG TTAGGG..3'
3'...AATCCC AATCCC AATCCC AATCCC AATCCC AATCCC..5'
64.
65. 65
From Principles of Genetics,
Snustad, Simmons and Jenkins
eds. John Wiley and Sons
Progeria nl.
Hutchinson-
Gilford syndrome
Premature ageing
correlated with
shorter telomeres
69. So sánh tổng hợp ở chân hạch và sơ hạch
1. Giống nhau:
1). Hai sợi poly của DNA cùng làm khuôn tổng hợp DNA
mới. Tổng hợp bắt đầu từ đầu 5’ đến đầu 3’của sợi pol
mới
2). Tổng hợp bắt đầu từ khởi điểm và tiến hành theo hai
hướng ngược chiều nhau tạo thành chán hai. Một pol làm
khuôn tổng hợp một sợi pol liên tục; sợi pol đối diện làm
khuôn tổng hợp nhiều đoạn poly không liên tục gọi là các
đoạn Okazaki
3). DNA polymeraz cần có sự hổ trợ của nhiều loại protein và
sự hiện diện của ARN prime để bắt đầu tổng hợp sợi pol.
70. 2. Khác nhau:
1). DNA của vi khuẩn chỉ có một khởi điểm; DNA của
tế bào chân hạch có nhiều khởi điểm
2). Vi khuẩn chỉ có một loại pol.III; chân hạch có hai
loại pol III( pol và pol)
3). Ở vi khuẩn dnaA là protein đầu tiên tương tác với
DNA trong khi ở chân hạch là protein T
4). DNA vòng kín ở vi khuẩn làm khuôn tổng hợp DNA
mới theo nguyên tắc bán bảo toàn; DNA tế bào chân hạch
làm khuôn tổng hợp pol mới có đầu 3’ mang nhiều đoạn
lập lại nhiều lần (telomer)????
So sánh tổng hợp ở chân hạch và sơ hạch
71.
72. Cơ chế tổng hợp DNA-
Cơ chế tổng hợp đặc biệt [Cơ chế tháo chiếu ở một số vi rút
(x174)]
DNA của x174 sợi đơn vòng kín
1. Sau khi vào tế bào vi khuẩn, DNA ( gọi là DNA dương-
pol.+) làm khuôn tổng hợp sợi âm cho ra DNA sợi đôi
vòng kín gồm hai vòng pol.+ và pol.-
2. Sợi pol.- làm khuôn tổng hợp pol. +: Vòng + bị enzim (do
vi rút chỉ đạo tổng hợp) cắt tại khởi điểm và tổng hợp
tiến hành ở đầu 3’ hydoxyl tự do và dùng Pol.- làm
khuôn. Khi sợi pol + tháo ra đủ dài, nó bị cắt và nối cho
ra phân tử DNA sợi đơn+ vòng kín của vi rút
75. Một cách khác để tạo ra DNA
RNA-Directed DNA Polymerase
1. Năm 1964: Howard Temin nhận thấy rằng
các chất ức chế tổng hợp DNA ngăn chặn
sự lây nhiễm của các tế bào trong quá
trình nuôi cấy bởi virus khối u RNA.
Temin dự đoán rằng DNA là chất trung
gian trong quá trình nhân lên của virus
khối u RNA
2. 1970: Temin và David Baltimore (độc lập)
khám phá ra “Reverse transcriptase"
76. Làm thế nào để các bộ gen RNA
được tái tạo?
1. Nhiều loại virus có bộ gen bao gồm RNA
2. DNA là chất trung gian trong quá trình sao
chép của virus RNA
3. RNA của virus đóng vai trò là khuôn mẫu để
tổng hợp DNA
4. RNA-directed DNA polymerase được gọi là
Reverse Transcriptase
77. Reverse Transcriptase (RT)
1. Cần có mồi nhưng phải có mồi lạ - phân
tử tRNA mà vi rút bắt được từ vật chủ
2. RT phiên mã khuôn mẫu ARN thành ADN
bổ sung (cDNA) để tạo thành phép lai
ADN: ARN
3. Tất cả các vi rút khối u RNA đều chứa
một enzyme phiên mã ngược
78. Hoạt động của Reverse
Transcriptase
1. Ba hoạt động của enzyme
2. RNA-directed DNA polymerase
a) Hoạt động của RNase H - phân hủy RNA
trong DNA: RNA lai
b) DNA-directed DNA polymerase - tạo ra
một DNA kép sau khi hoạt động của
RNase H sẽ phá hủy bộ gen của virus
c) Điều trị HIV: AZT (hoặc 3'-azido-2 ', 3'-
dideoxythymidine) ức chế đặc hiệu RT
79.
80.
81. Các tế bào đang sống (bao
gồm những loại đơn giản nhất)
Làm đúng lại chức năng của gen
Đột biến
Đột biến gen Đột biến nst
Có hai loại đột biến: Đột biến tự phát và đột biến do bị
kích thích
84. Các cơ chế sửa của DNA
Cắt bỏ baz:
Các baz trong phân tử DNA co thể bị biến đổi bởi sự
loại bỏ gốc amin hay alkyl hoá. Vị trí của baz bị hư
hỏng gọi là vị trí AP (abasic position). Ở E.coli, DNA
glycosylase có thể nhận biết được vị trí AP và cắt bỏ
baz. Các nucleosid sau khi bị loại bỏ baz hư sẽ được
endonuclease loại bỏ AP và cắt bỏ thêm một số các
nucleotid nằm cạnh. DNA polymerase I sẽ bổ sung lại
những baz đúng tại các vi trí đó, DNA ligase giúp hàn
gắn lại mạch DNA bị cắt đứt
85. Figure 7-G-1. DNA repair by base excision
Cắt bỏ baz sai
Cắt bỏ các nucleosis có
baz sai
86. Cắt bỏ Nucleotide
Ở E. coli, các proteins UvrA, UvrB, and UvrC có
chức năng loại bỏ các nucleotid bị hư hỏng. Thí dụ
tia UV. (hình thành cấu trúc dimer như thymin
dimer). DNA polymerase I sẽ bổ sung lại những baz
đúng tại các vi trí đó, DNA ligase giúp hàn gắn lại
mạch DNA bị cắt đứt.
Ở nấm men, các protein có chức năng giống các
protein Uvr có tên là RAD (RAD là từ viết tắt của
bức xạ radiation). Thí dụ như RAD3, RAD10
Các cơ chế sửa sai của DNA
87. Dưới tác đông của tia tử ngoại, thymin bị đứt liên kết C=C mạch vòng
nối 2 pt lại thành Thymin dimer
88.
89. Sửa chữa các baz bắt cặp sai (Mismatch repair)
• Xảy ra trong quá trình sao chép DNA
• Methy hoá các baz Adenin A tại các trình tự GATC
ở đầu 5’ của sợi cũ (sợi khuôn) bởi enzyme dam
methylase.
Các cơ chế sửa sai của DNA