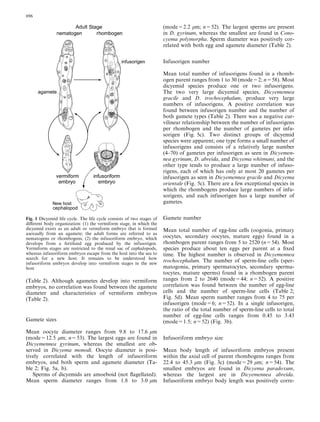
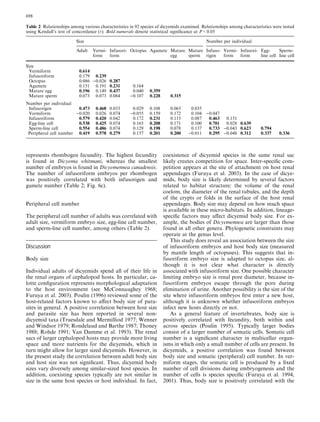
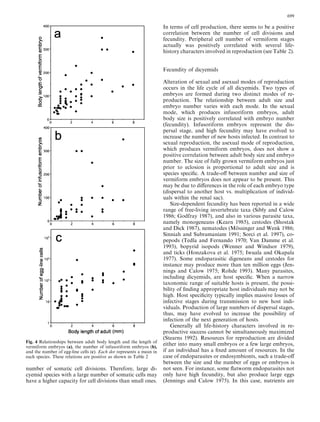
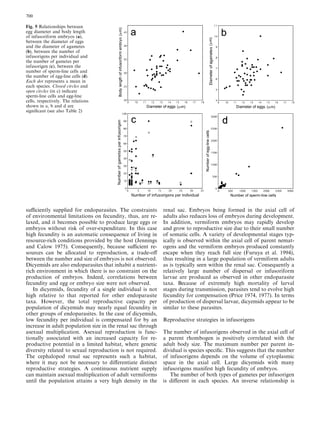
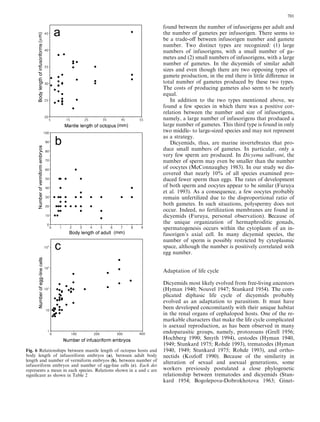
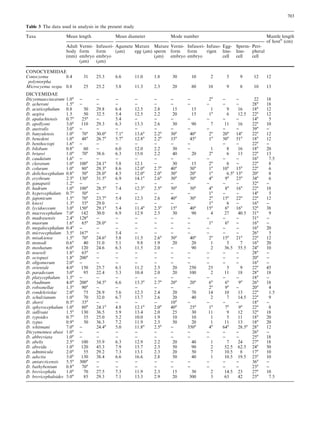
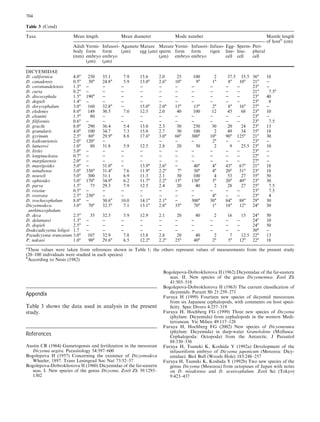
This document examines various life history traits of dicyemids to understand their reproductive strategies and adaptations to living as endoparasites in cephalopod renal organs. It finds that dicyemids have a hermaphroditic gonad that produces roughly equal numbers of eggs and sperm. Fecundity increases with adult body size. Embryo size correlates with host size, suggesting host factors influence dispersal and infection of new hosts. While individual fecundity is low, total reproductive capacity per community is high due to asexual multiplication within the host. Adult size appears constrained by the volume and features of the renal habitat within different host species.