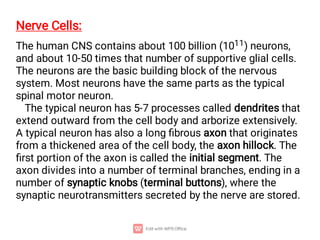
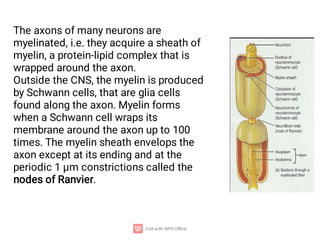
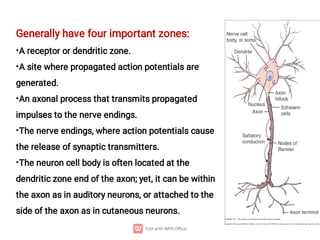
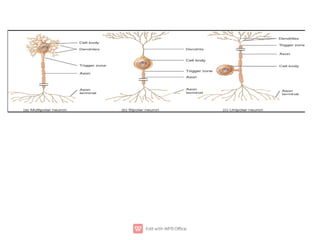
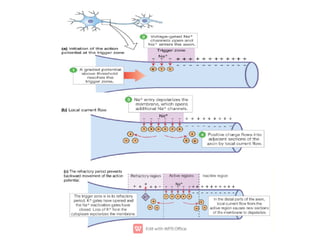
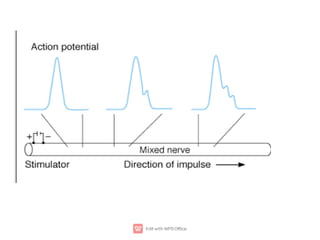
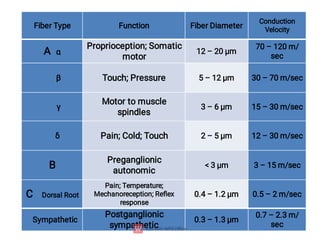
1. Nerve and muscle cells are excitable tissues that can generate electrochemical impulses. In nerves, these impulses propagate signals along the axon when stimulated, whereas in muscles they cause contraction.
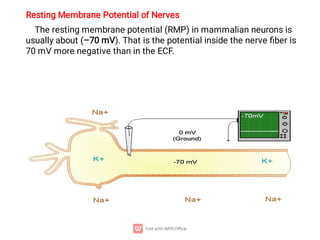
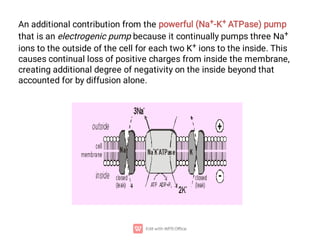
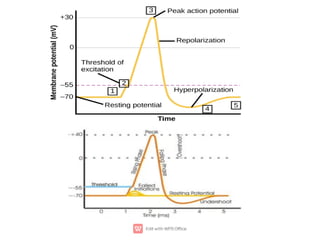
2. The resting membrane potential of these cells is maintained by ion concentration gradients and selective permeability of the membrane to ions like sodium, potassium, and chloride. At rest, the intracellular fluid is negatively charged relative to the extracellular fluid.
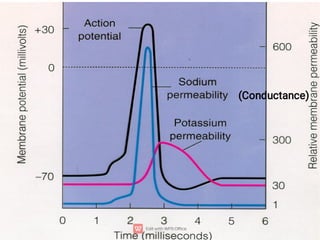
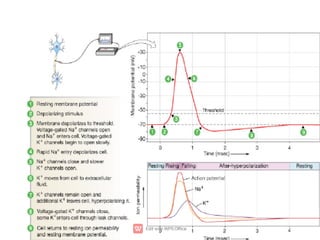
3. An action potential occurs when the membrane potential rapidly changes from the resting potential to a positive overshoot then back again. This is driven by changes in sodium and potassium conductance across the membrane.





![Nernst Equation:
Equilibrium Potential (E):
E = + 61 X log
Ion concentration inside
Ion concentration outside
- 61
EK
+ = X log
[ K+ ] inside
[ K+ ] outside
= - 94 mV
X log [ Na+ ] inside
[ Na+ ] outside
= + 61 mV
ENa
+ = - 61](https://image.slidesharecdn.com/nerveandmuscle1-230502153327-66328396/85/Nerve-and-Muscle-1-pdf-6-320.jpg)