Downloaded 94 times



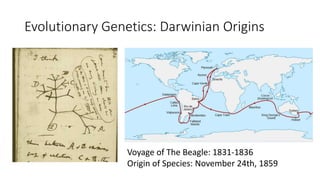


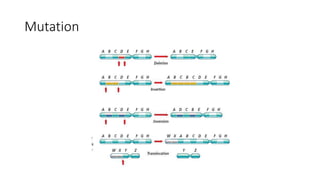
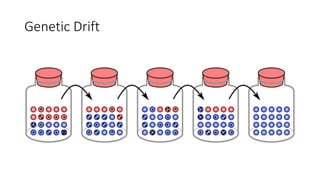






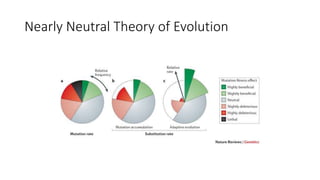

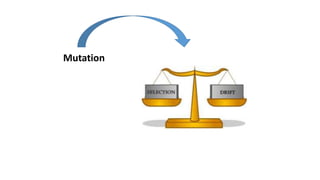

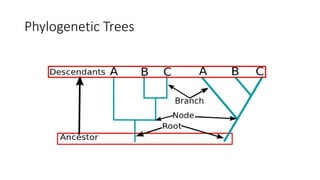





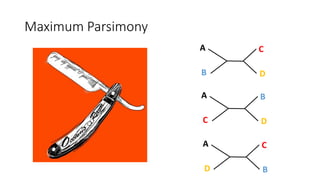
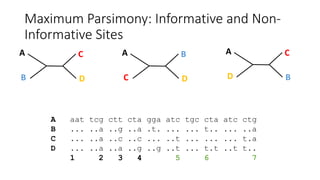
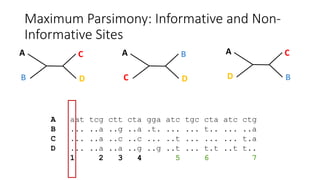
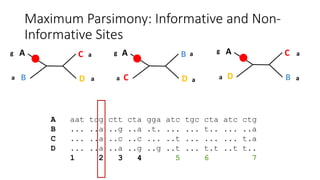
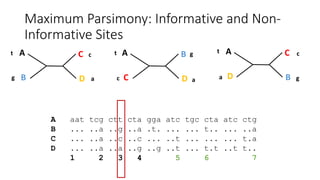
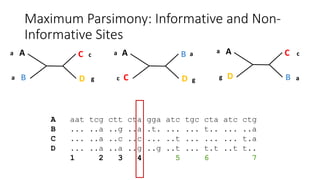
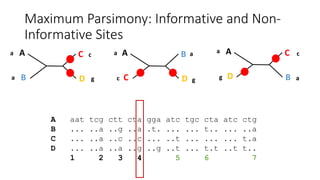
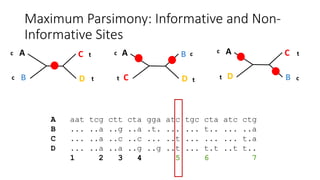



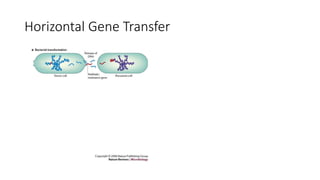
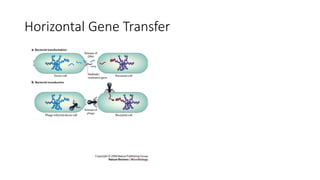
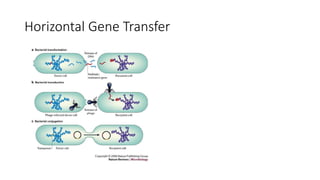
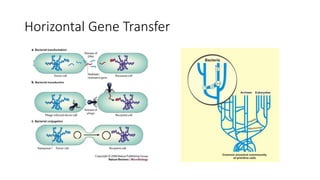
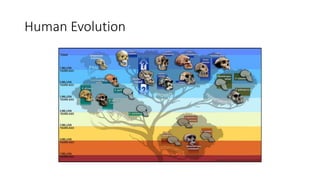
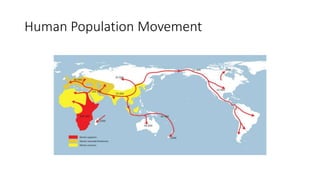
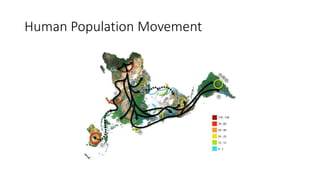
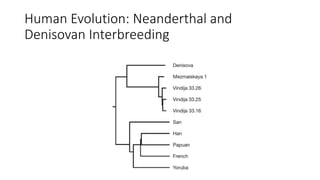
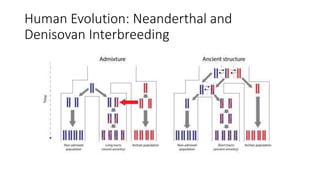




1) Evolutionary genetics examines how genetic changes in populations lead to evolution over time through processes like mutation, genetic drift, and natural selection. 2) Early work in evolutionary genetics integrated Mendelian genetics with Darwin's theory of evolution through natural selection to form the modern synthesis of the early 20th century. 3) Neutral theory proposed that many genetic mutations are neutral or nearly neutral and do not significantly impact fitness, explaining much molecular evolution. 4) Phylogenetic trees reconstructed from genetic data provide evidence for relationships between species and models of molecular evolution. 5) Horizontal gene transfer and archaic human interbreeding have introduced genetic variation influencing traits like placental invasiveness and immunity.












































































![Polymer [ बहुलक ] Chemistry Notes PDF - Irfanullah Mehar - JJ Sir Chemistry.pdf](https://cdn.slidesharecdn.com/ss_thumbnails/polymerchemistrynotespdf-irfanullahmehar-jjsirchemistry-260210172118-3f9b37f7-thumbnail.jpg?width=640&height=640&fit=bounds)

