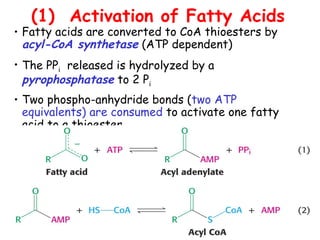
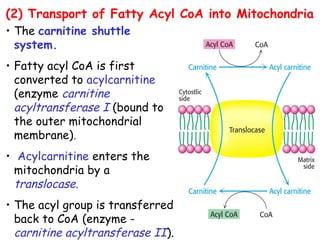
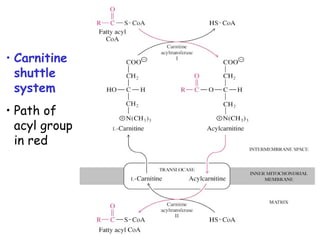
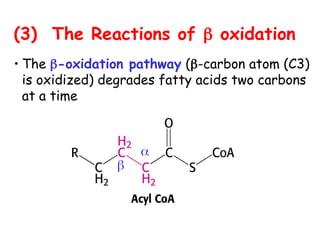
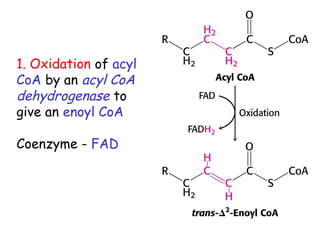
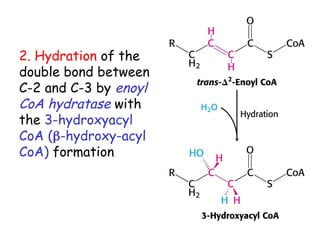
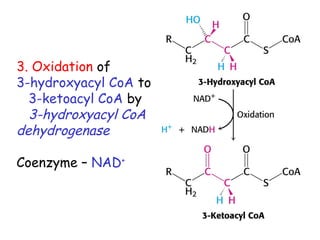
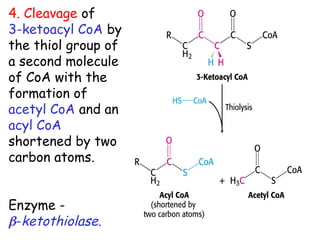
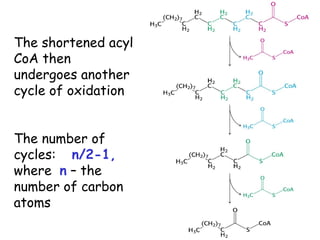
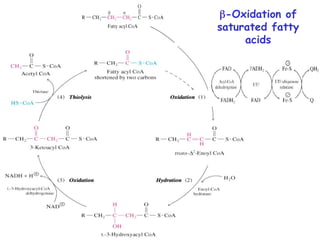
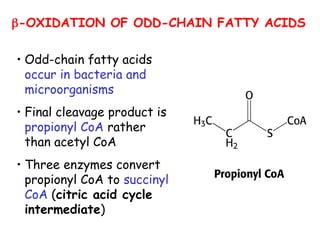
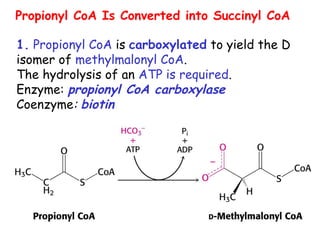
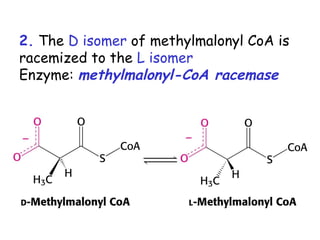
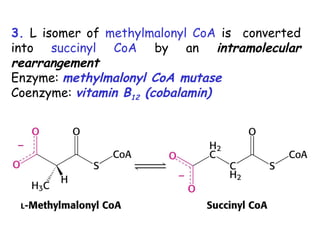
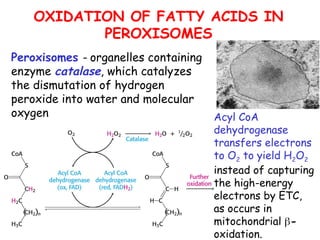
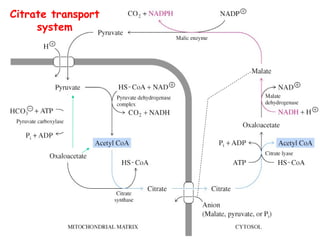
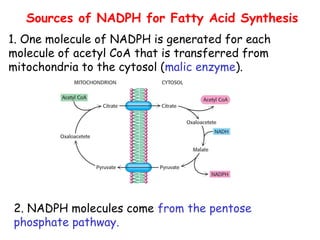
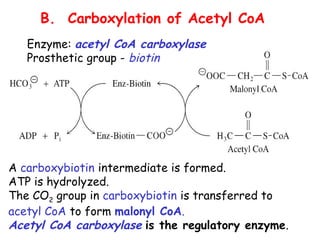
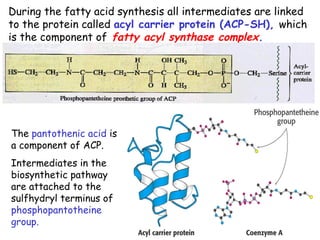
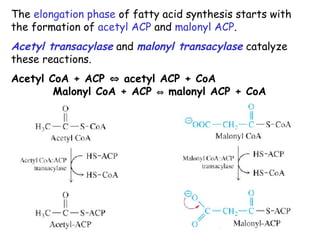
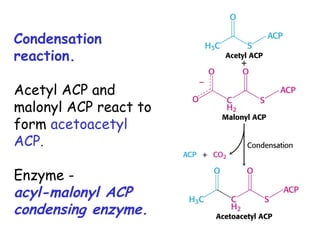
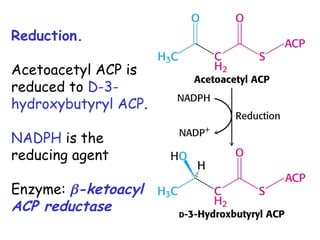
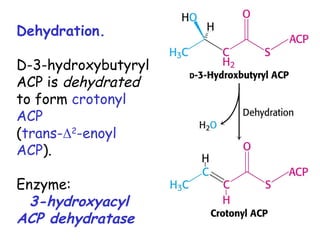
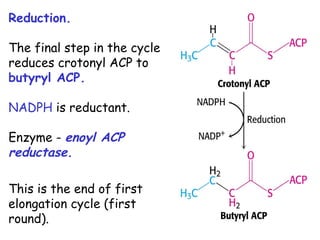
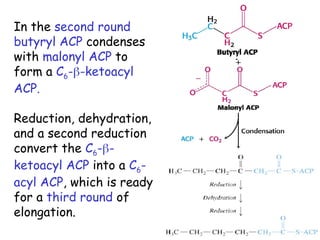
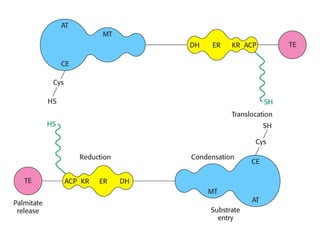
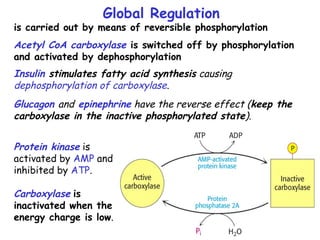
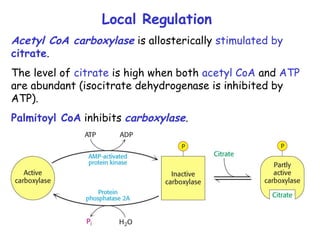
The document provides a comprehensive overview of fatty acid metabolism, detailing the processes of fatty acid oxidation in mitochondria and peroxisomes, as well as fatty acid synthesis primarily occurring in the cytoplasm. It covers the enzymatic pathways involved, including the carnitine shuttle system, beta-oxidation, and the synthesis stages from acetyl-CoA, highlighting the energy yields and regulatory mechanisms, such as the role of acetyl-CoA carboxylase and hormonal influence. Additionally, it explains the importance of these pathways in energy production and metabolic control in response to dietary intake.