Download to read offline

















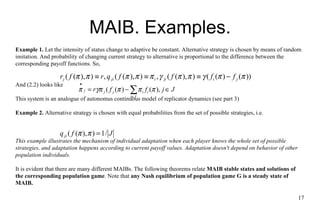









































This document discusses concepts from evolutionary game theory and economics. It summarizes key principles like Nash equilibrium, dominance elimination, and evolutionary stable strategies. It also provides an example of a population game modeling pair contests over resources, and discusses how static optimality principles like Nash equilibrium apply in this context. Dominance and evolutionary stable strategies are presented as alternative criteria when multiple equilibria exist.






















































































