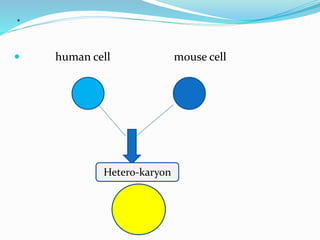
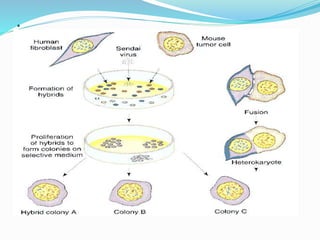
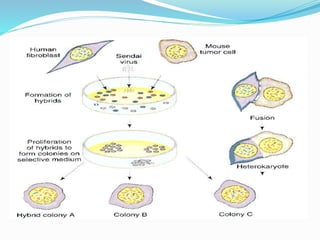
Somatic cell hybridization involves fusing cells from two different species, such as human and mouse cells, to form hybrid cells containing chromosomes from both species. This technique allows genes to be mapped to specific chromosomes. It works by using selective growth conditions that require the hybrid cell to retain certain human chromosomes in order to survive. Over successive cell divisions, human chromosomes are eliminated at random except for those required for survival. This allows the creation of cell lines containing partial sets of human chromosomes that can be analyzed to correlate genes with specific chromosomes. The technique has been important for mapping the human genome.