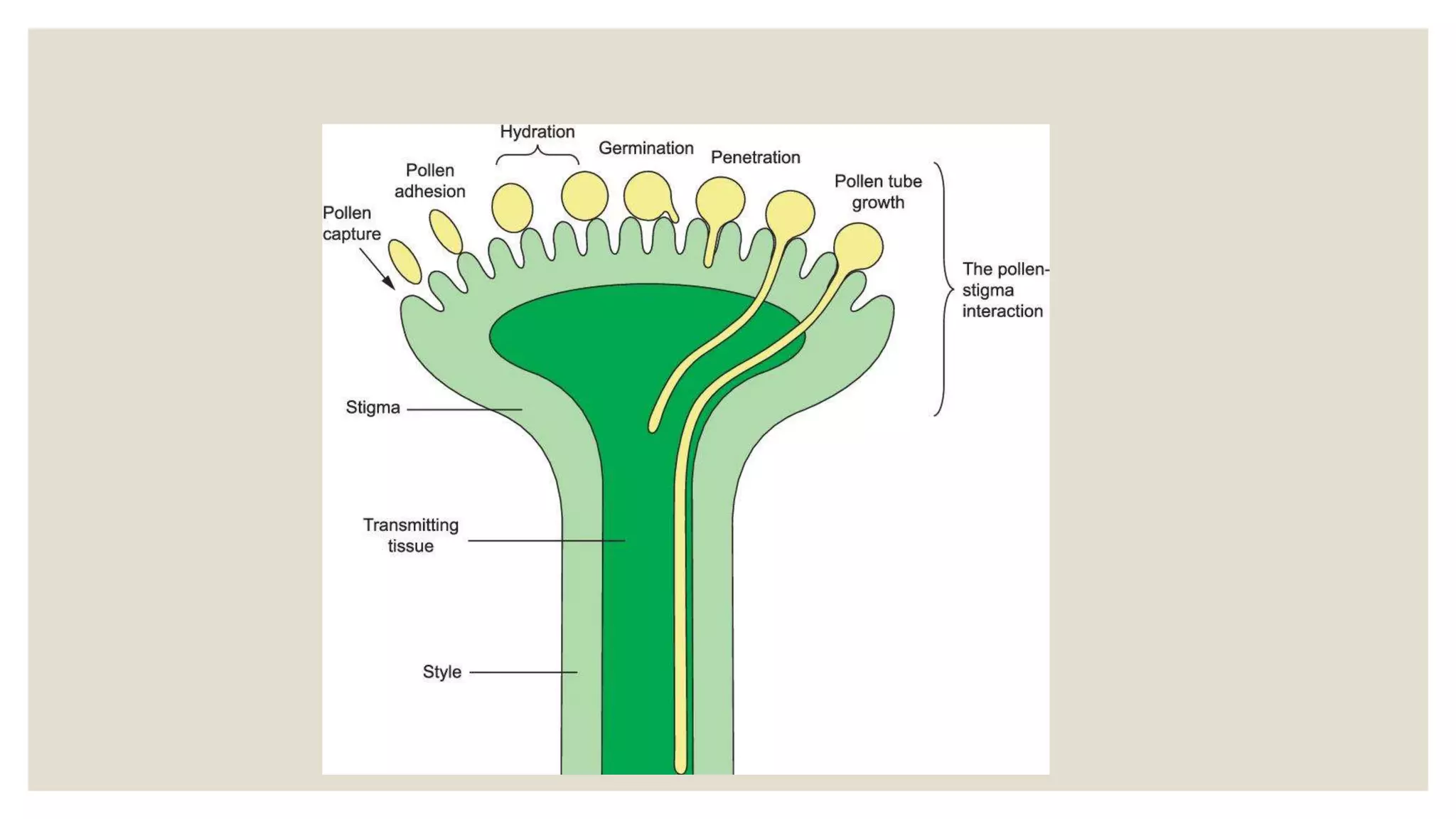
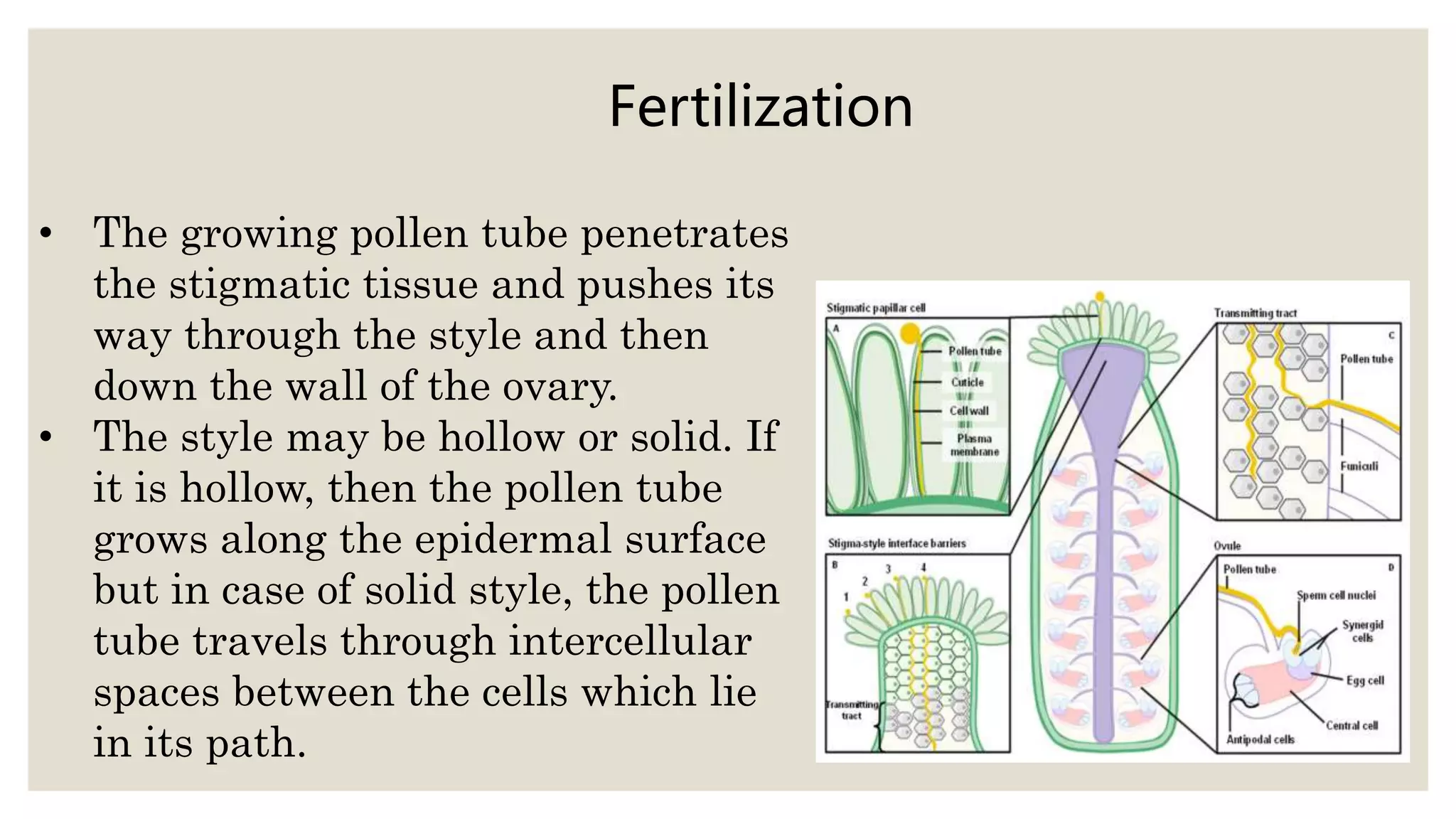
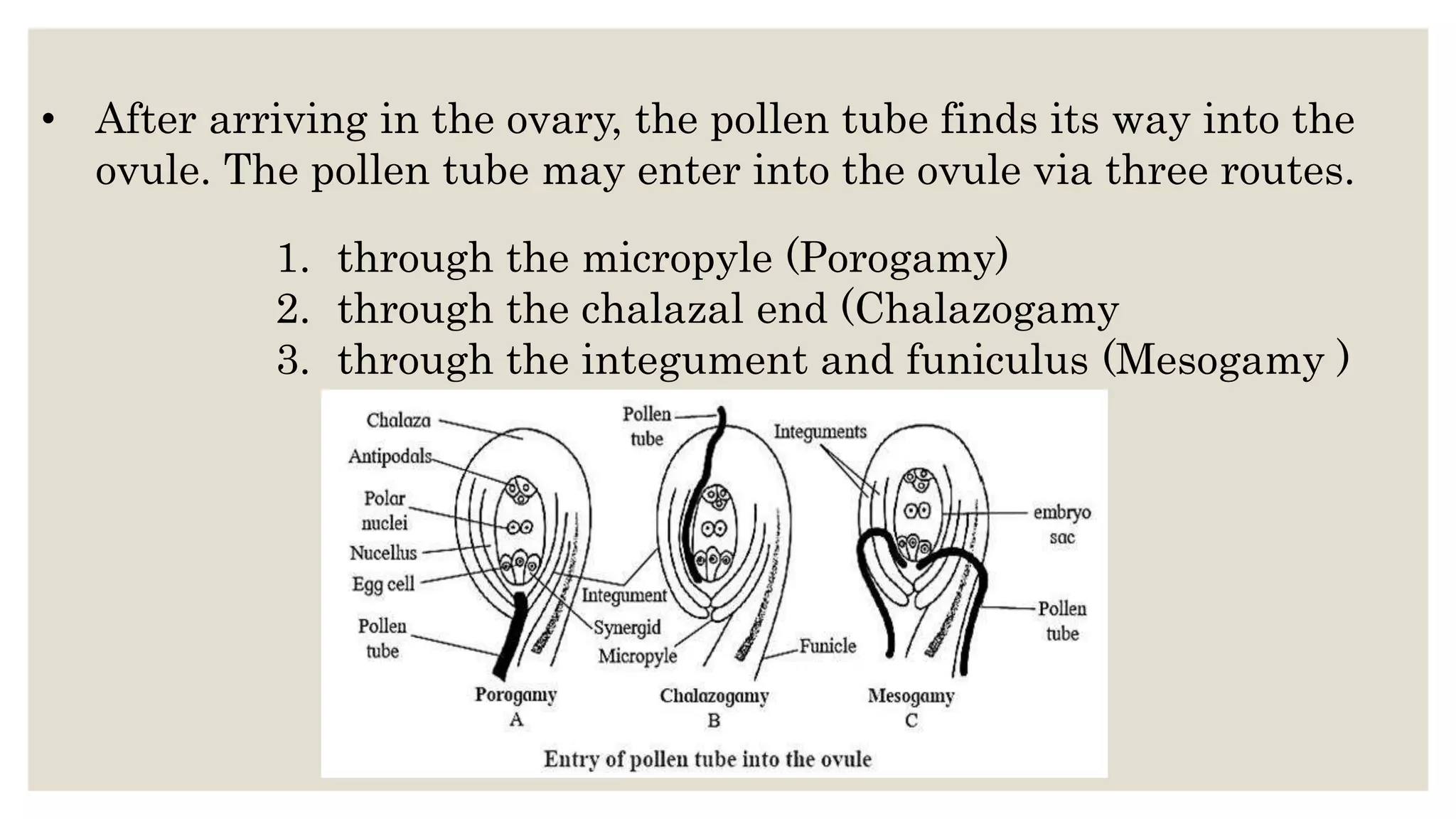
Pollen grains have a complex wall structure that protects them as they are carried by wind or animals to the stigma of another flower. Stigmas come in dry and wet types, with wet stigmas secreting fluids that hydrate pollen grains. Compatible pollen grains germinate and send tubes through the stigma and style to the ovary. Self-incompatibility prevents self-fertilization via heteromorphic or homomorphic floral structures. Double fertilization occurs when one sperm cell fuses with the egg to form the zygote, and the other with the polar nuclei to form the endosperm.