Download as PDF, PPTX











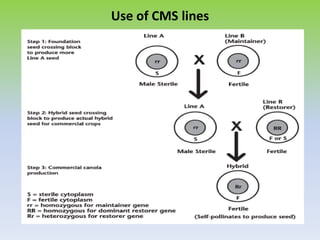
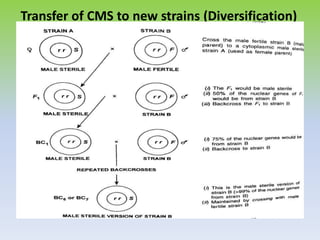



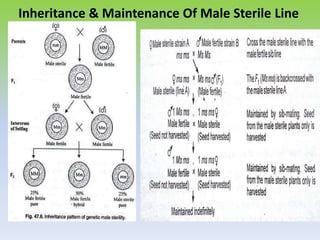
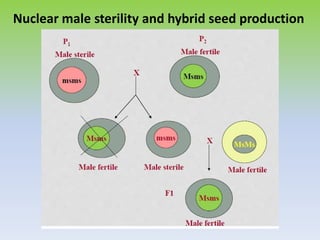

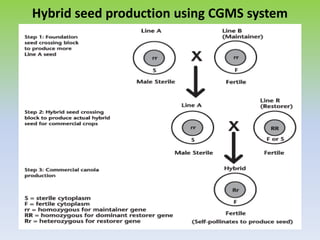
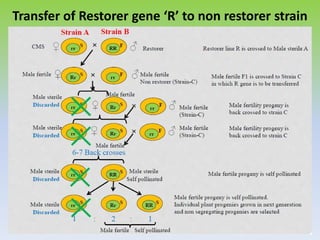
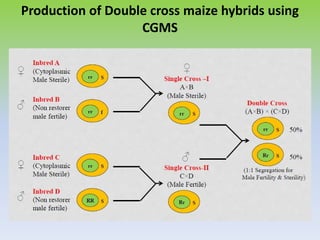
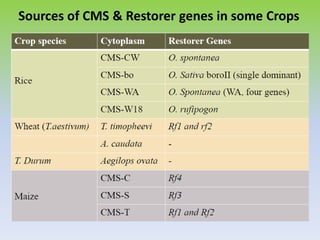
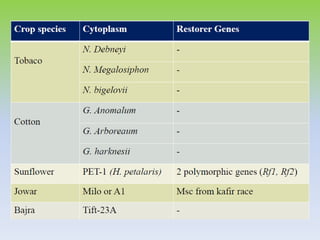
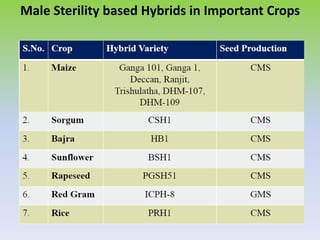


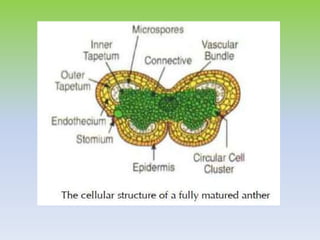

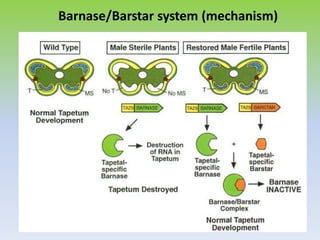
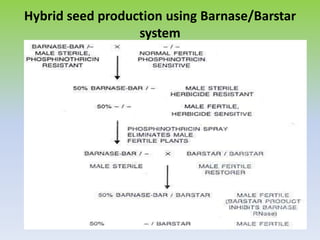










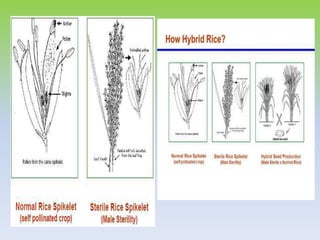
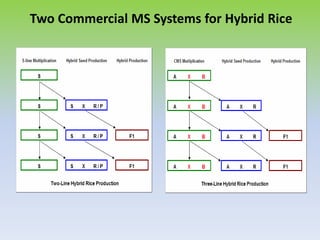
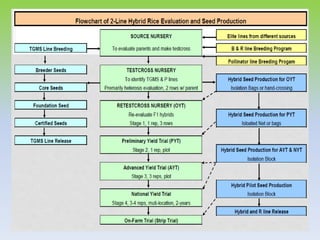
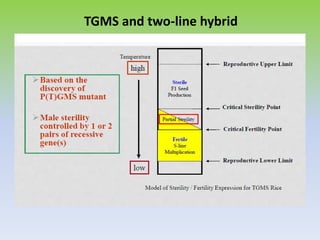


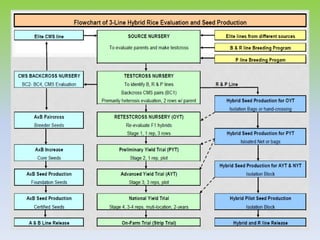

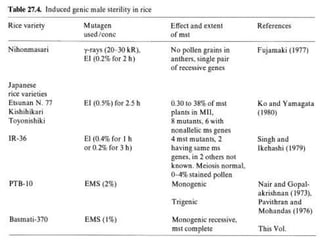
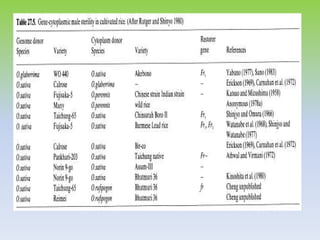

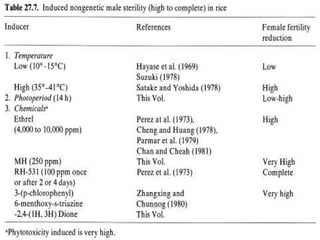
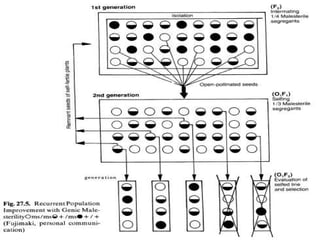
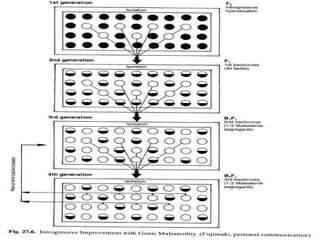
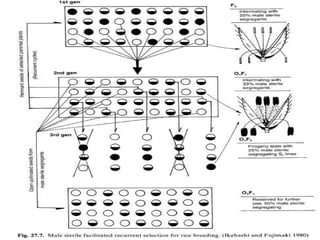
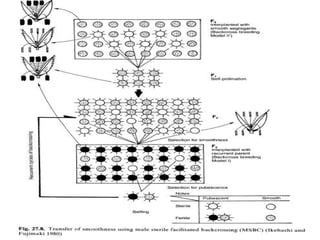
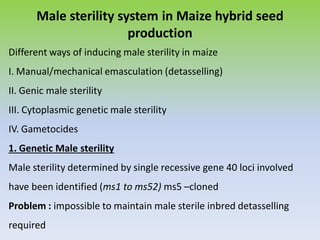

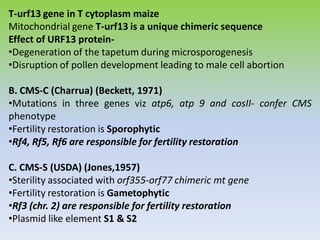
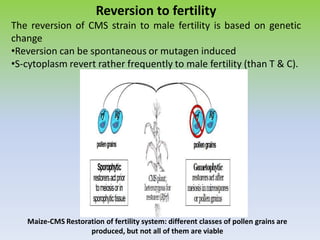

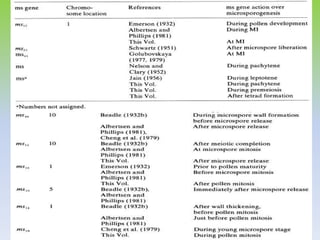
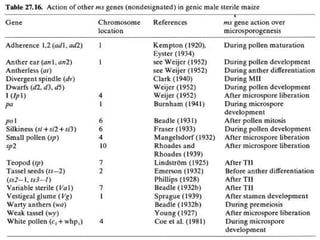
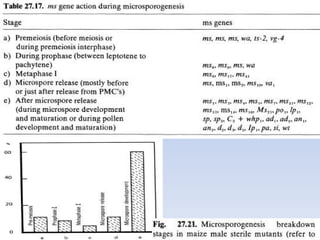
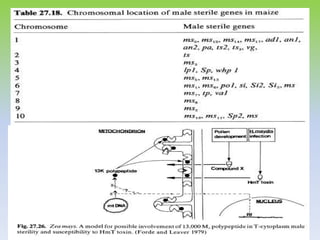
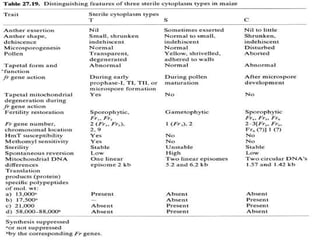
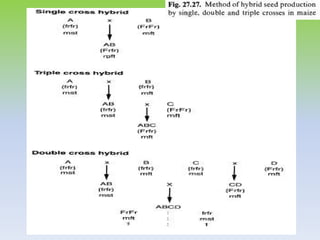



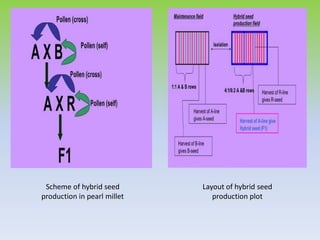


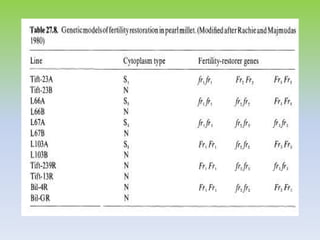
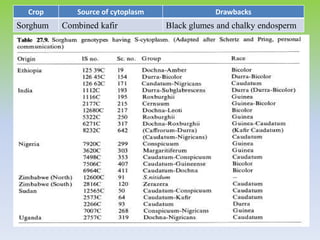




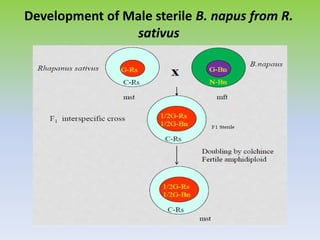
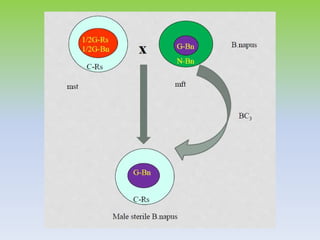
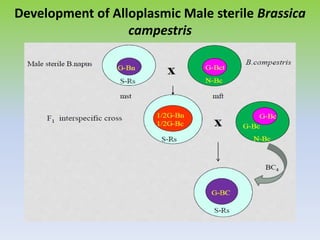
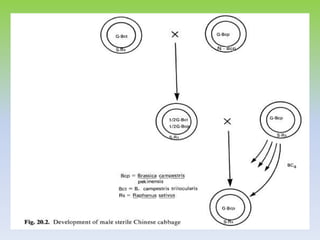
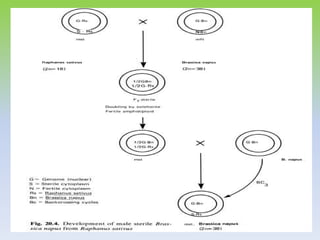



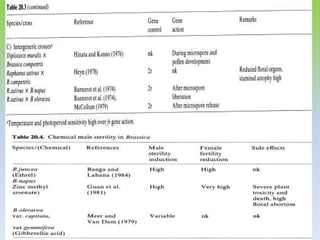
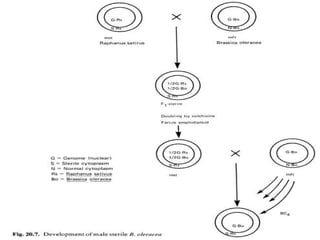






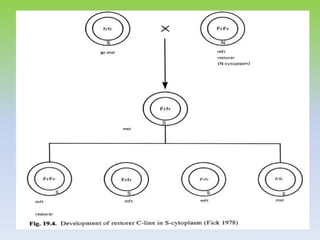
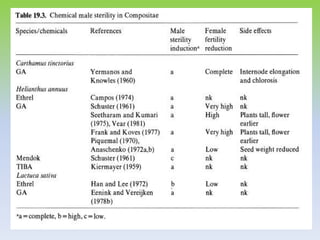

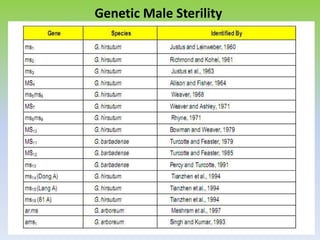
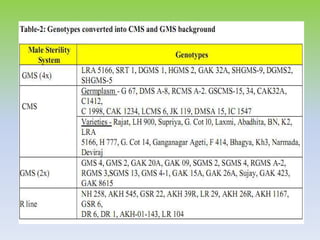




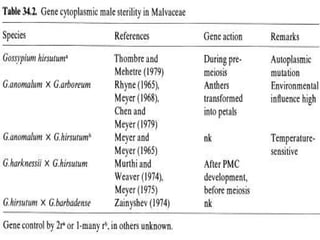
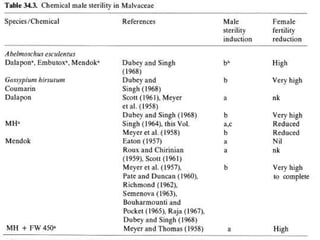


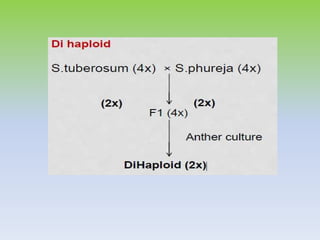




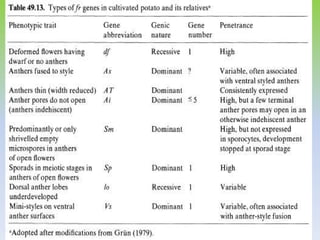
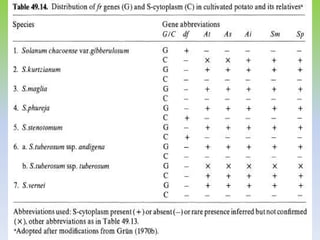

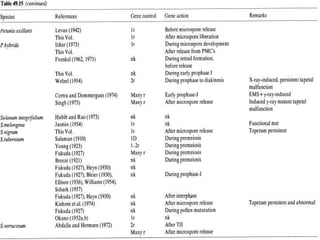





This document discusses different types of male sterility in plants, including genetic male sterility (GMS), cytoplasmic male sterility (CMS), and chemically-induced male sterility (CHA). It describes how each type of male sterility works and how it can be used for hybrid seed production. Specifically, CMS uses cytoplasmic genes to induce sterility and requires maintainer and restorer lines, while GMS uses nuclear genes and can be environmentally sensitive. The document also covers transgenic systems like Barnase/Barstar and provides examples of major crops where male sterility systems have been applied.























































































