

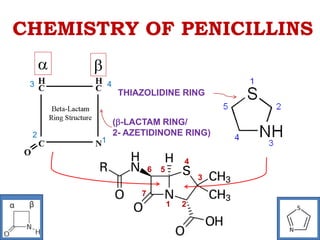
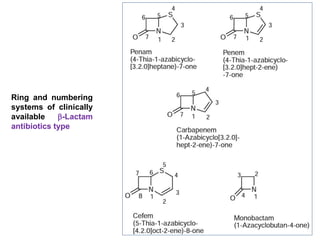
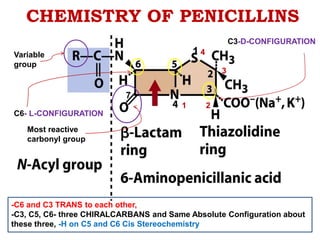
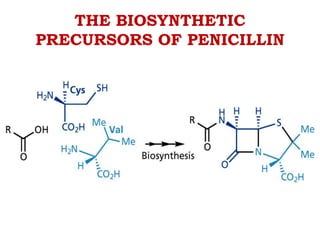
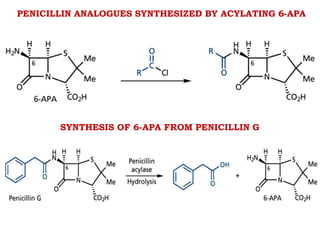
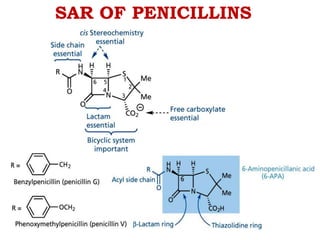
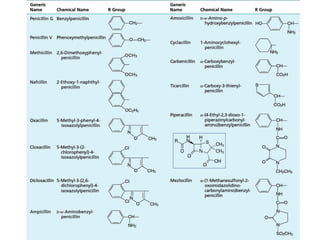
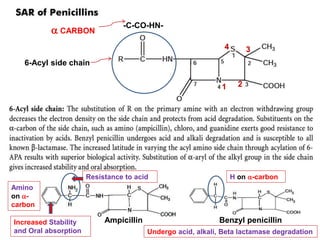
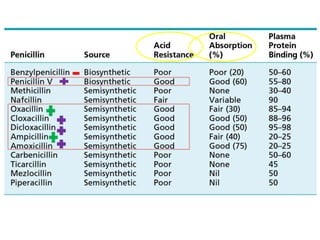
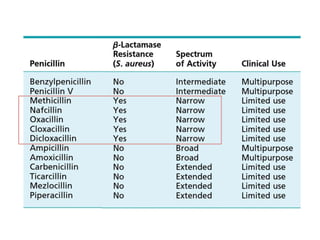
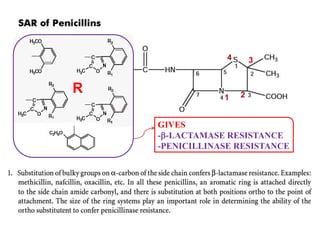
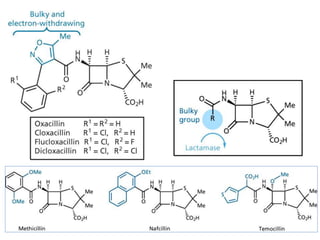
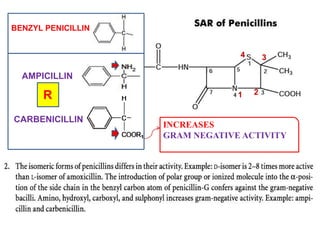
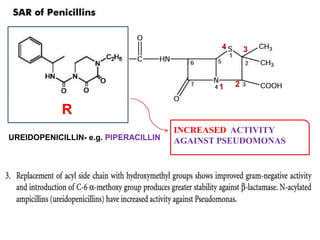
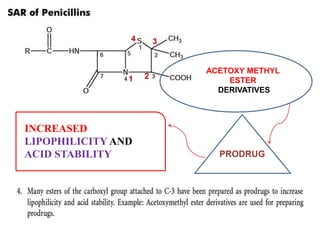

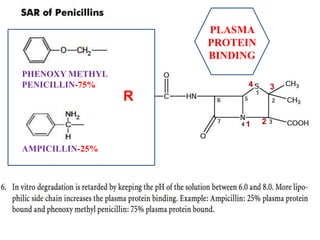
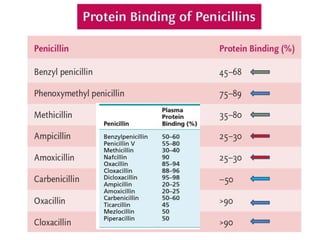
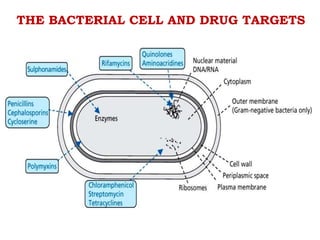

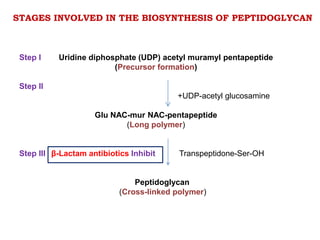
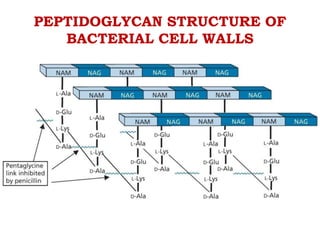
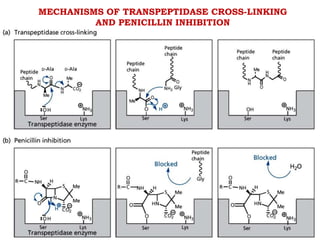
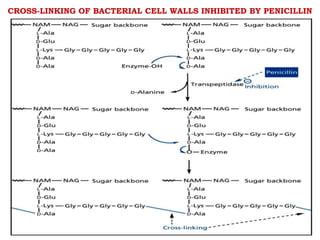
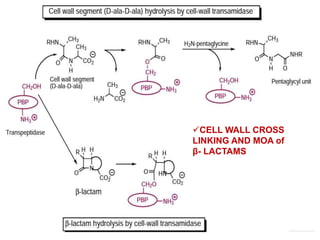
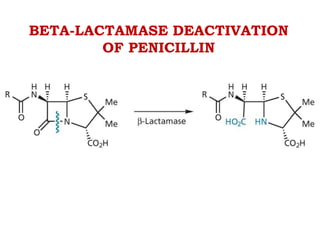






The document discusses the chemistry, structure-activity relationships (SAR), and mechanism of action (MOA) of penicillins, highlighting the β-lactam ring's role in inhibiting bacterial cell wall synthesis. It details how penicillins target transpeptidase enzymes, leading to defects in peptidoglycan cross-linking, which ultimately causes bacterial cell lysis. The interaction between penicillins and bacterial enzymes is emphasized, showing how penicillins mimic natural substrates, resulting in irreversible inhibition.















![ANTI-TB AND ANTI LEPROTIC DRUGS [MEDICINAL CHEMISTRY] BY P.RAVISANKAR.](https://cdn.slidesharecdn.com/ss_thumbnails/anti-tbdrugs-130615221719-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)
















































![ONFH[AVN HIP] -TRIPLE REGIME -A NOVAL SURGICAL CONCEPT .pptx](https://cdn.slidesharecdn.com/ss_thumbnails/onfhavnhip2026koaconcalicutdrgokuldevdrmashraf-260210064517-213ec005-thumbnail.jpg?width=640&height=640&fit=bounds)



![PERI-PROSTHETIC FRACTURE NAIL-PLATE CONSTRUCT [NPC].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/drarunkumardrmohamedashrafperiprostheticfrasturenail-plateconstructnpc-260209164459-7e9d15a1-thumbnail.jpg?width=640&height=640&fit=bounds)






