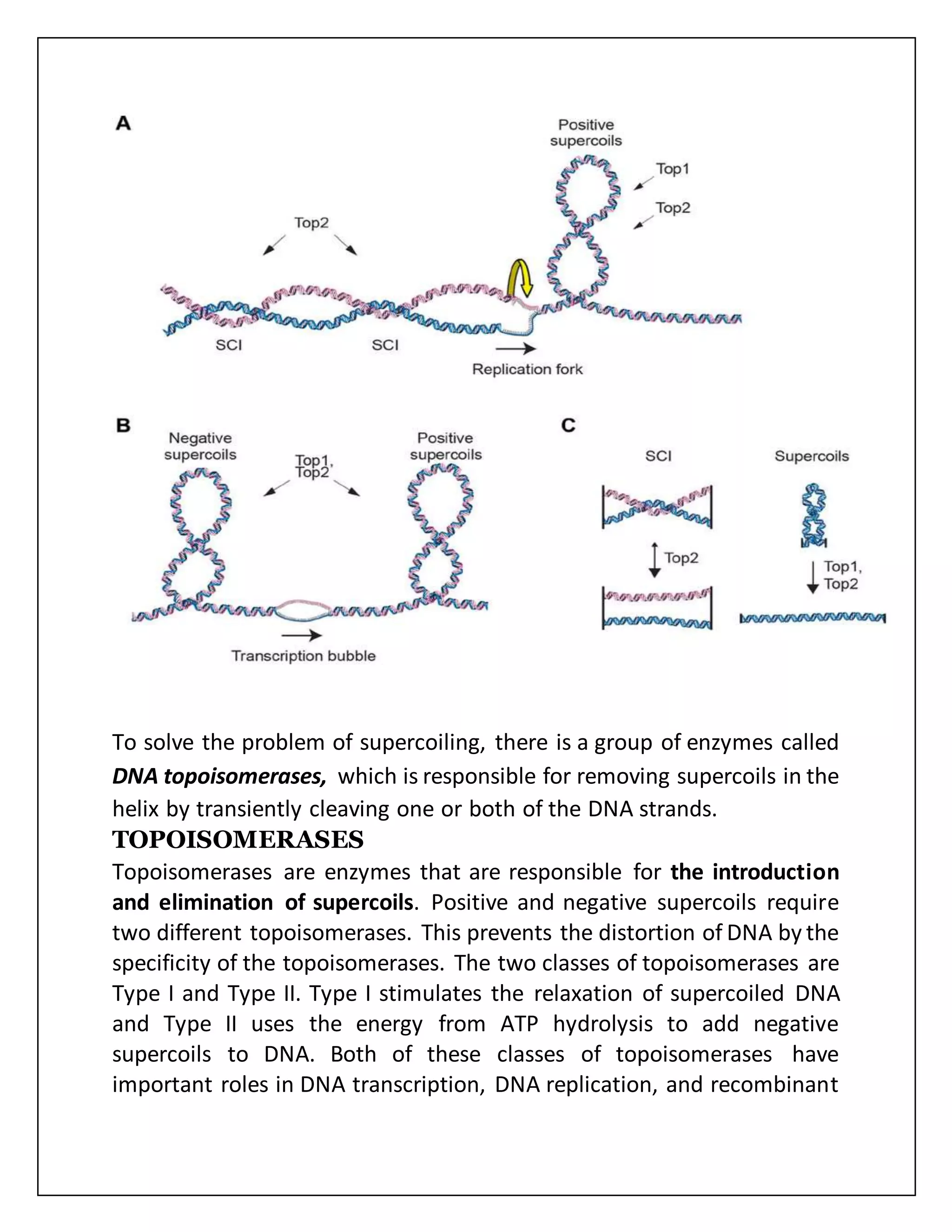
The document discusses the concept of DNA supercoiling, explaining how DNA molecules can exist in various forms, including relaxed and supercoiled states, and the impact of twisting on DNA structure and energy. It highlights the roles of topoisomerases in managing supercoils, with Type I enzymes relaxing supercoiled DNA and Type II enzymes capable of introducing negative supercoils using ATP. Supercoiling is crucial for processes like DNA replication and transcription, with negative supercoiling being particularly prevalent in prokaryotes and eukaryotes, as it facilitates strand separation.