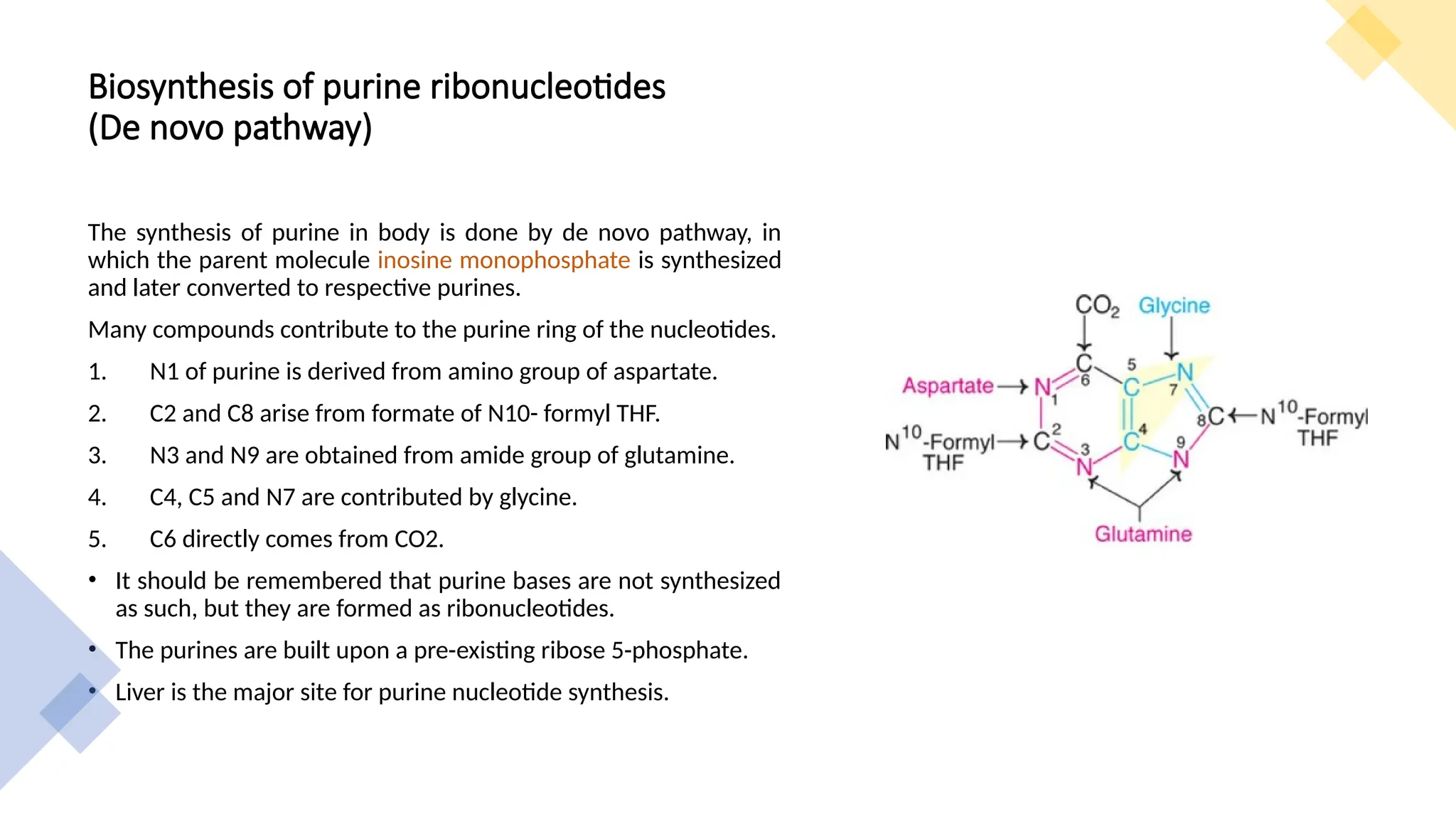
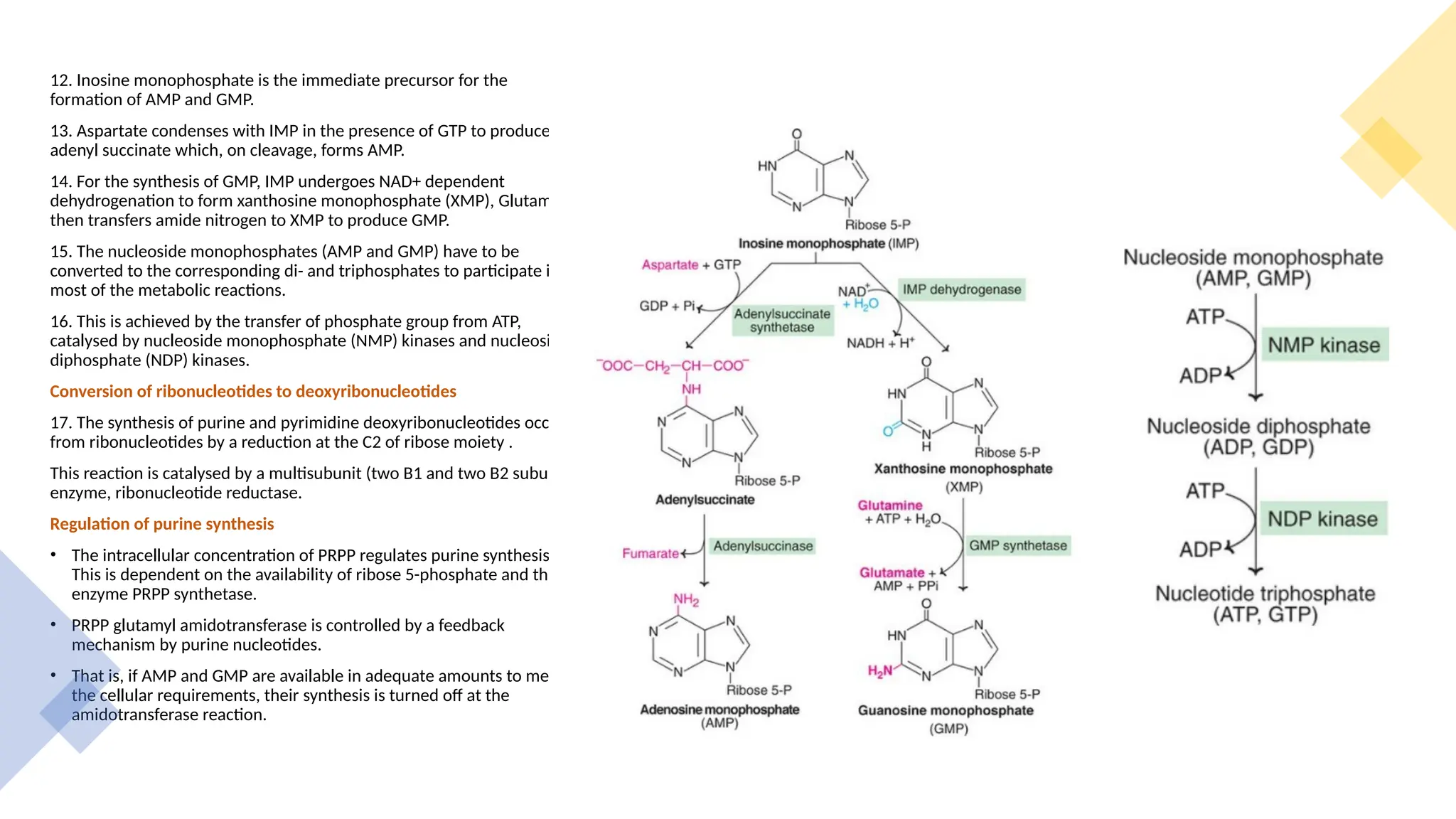
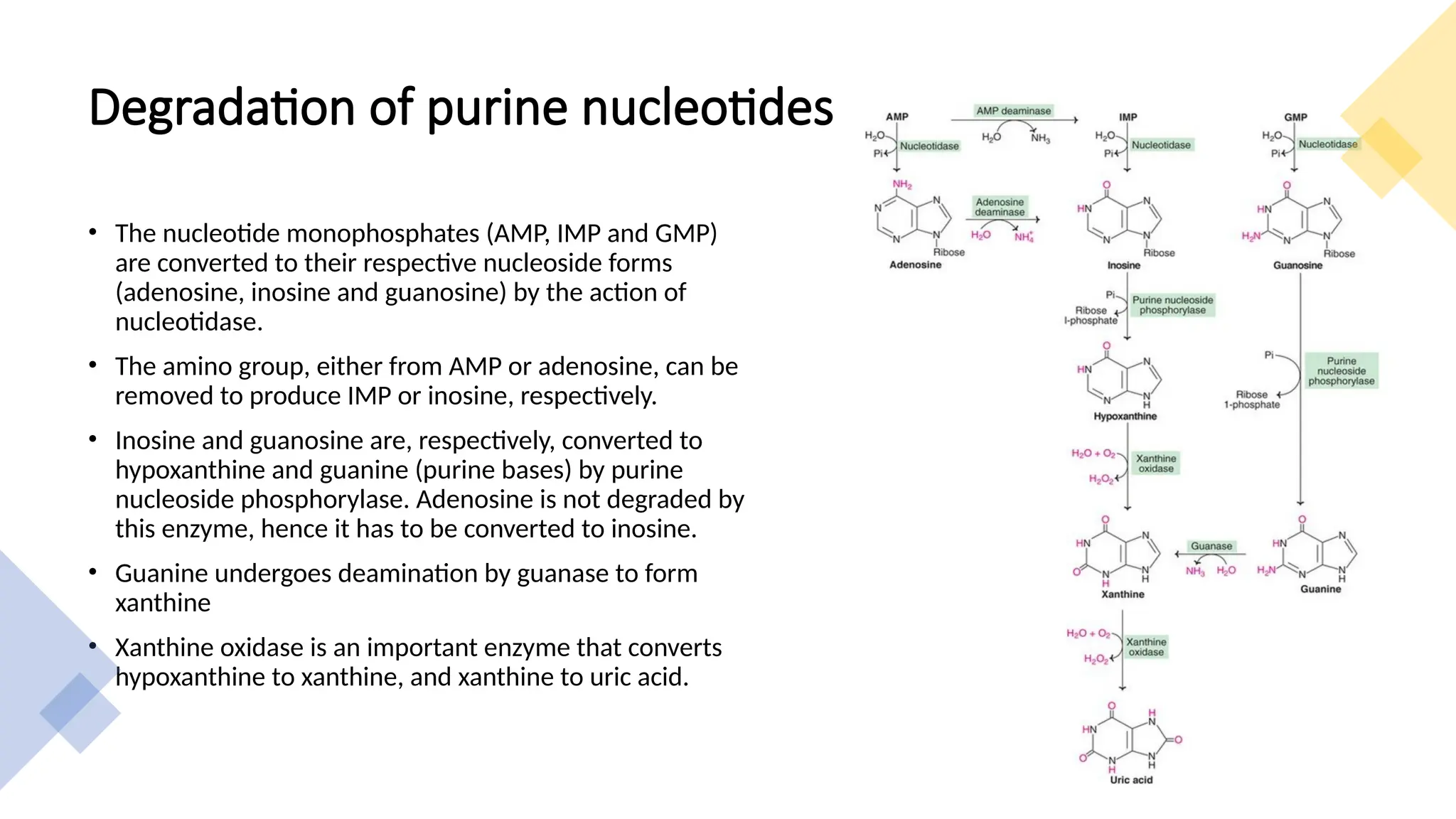
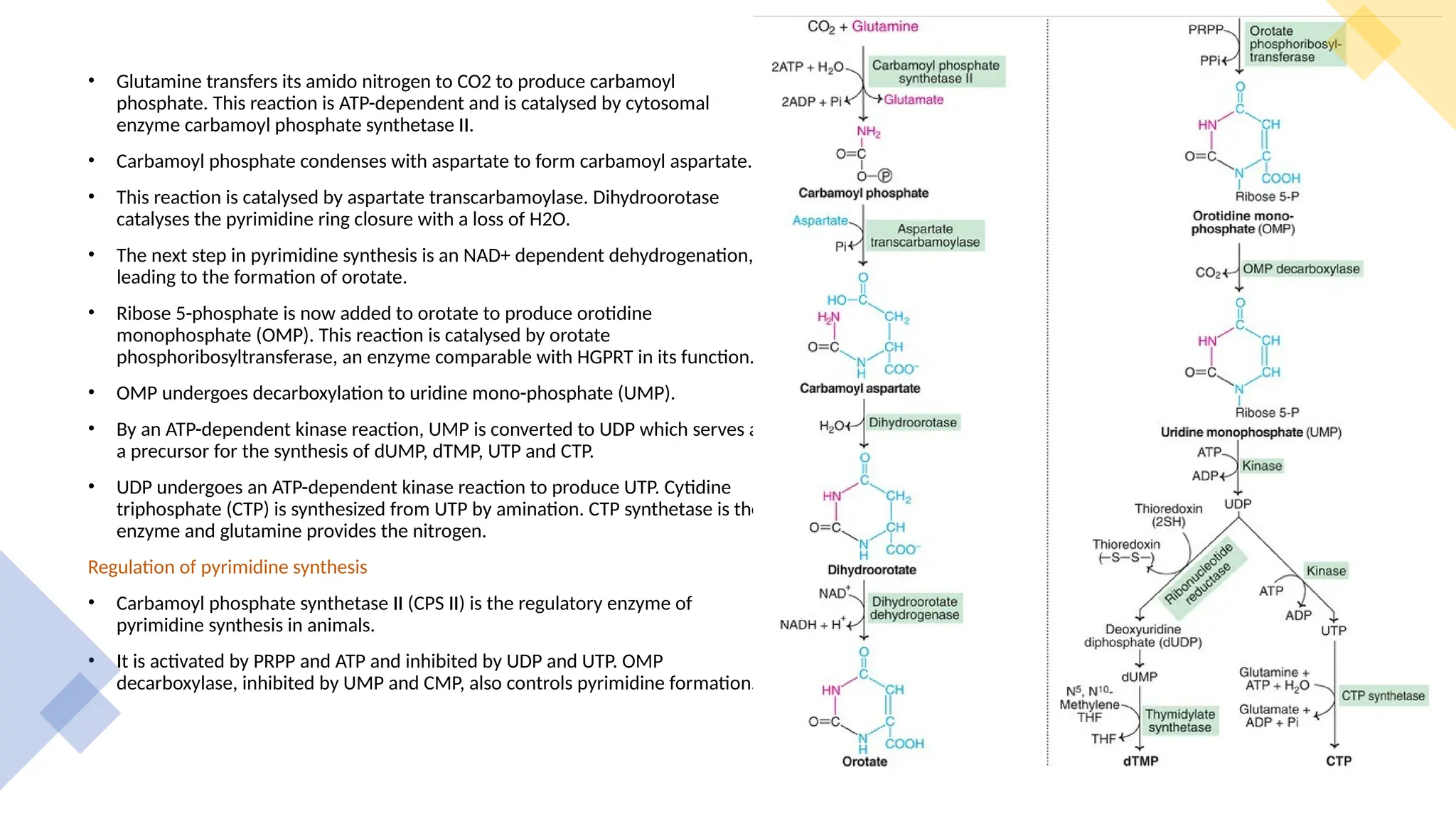
The document presents a detailed overview of nucleic acid metabolism, specifically focusing on the synthesis and degradation of purine and pyrimidine nucleotides. It describes the biosynthesis pathways, the roles of various enzymes, regulatory mechanisms, and the salvage pathways for nucleotide synthesis. Additionally, it outlines the degradation processes of purines and pyrimidines, leading to the formation of soluble products that are further metabolized.