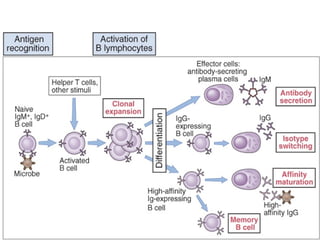








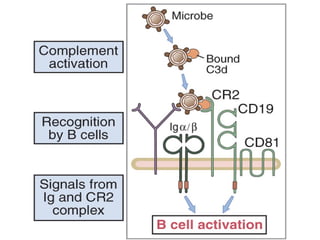

































- Naive B cells express IgM and IgD antibodies on their surface that recognize antigens. Upon activation, a single B cell can produce up to 10^12 antibody molecules per day through plasma cell differentiation. - Repeated antigen exposure leads to affinity maturation through somatic hypermutation in germinal centers, increasing antibody affinity over time. Helper T cells are required for isotype switching and affinity maturation. - Engagement of complement receptors and toll-like receptors enhances B cell activation and antibody production. Activated B cells also upregulate costimulators to amplify helper T cell responses.