Demir Metabolizması
Demir, birkaçbakteri türü hariç tüm canlı organizmalar için
“esansiyel” bir elementtir. Demir insanlarda eritropoetik fonksiyon,
oksidatif metabolizma ve hücresel immunite için gerekli olması
nedeniyle oldukça önemli bir elementtir. Elektron alıp verme özelliği
nedeniyle oksijen taşınması, enerji üretimi, DNA, RNA ve protein
sentezinde yer almaktadır. Birçok enzimin yapısına katılır ve/veya
fonksiyonu için gereklidir.
Erişkin bir erkekiçin vücuttaki toplam demir miktarı, 3500 mg’dır
(50 mg/kg). Vücuttaki demirin çoğu, hemoglobinler içinde dağılım
gösterir (% 65-70; 2300 mg). Yaklaşık olarak % 10’u (350 mg) kas lifleri
içinde (miyoglobin) ve diğer dokulardadır (enzimler ve sitokromlar).
Kalan %20-30’luk demir ise karaciğerde (200mg), retiküloendotelyal
sistem makrofajlarında; (500mg) ve kemik iliğinde (150mg)
depolanmaktadır. (Demirhavuzuoldukça değişken olup, hepatik demir;
reprodüktif çağdaki kadınlarda yaklaşık 300 mg., erişkin erkeklerde
yaklaşık 1 gr. kadardır.)
Vücuttaki miktarı, intestinal emilim ile düzenlenen bu elementin
fazlası toksik olup, fazla demirin atılımı ile ilgili bir mekanizma
bulunmamaktadır. Kanamalarla, gastrointestinal sistemden dökülen
epitelial hücrelerle, aşırı miktarda demir birikimi olan hastalarda ise
demir yüklümakrofajların dökülmesiyle oluşan demir kaybından başka
demir atılım mekanizması bulunmamaktadır.( Bu durum aşırı demir
2.
birikiminin altında yatangerçeğin, demirin intestinal mukozadan
emilimindekibozukluk olduğunu göstermektedir.)(şekil 1)
Şekil 1
Demir, insan vücudunda daima iki oksidasyon durumu olan ferrik
(Fe+3) veya ferröz (Fe+2) şekilde bulunur. Demirin bu elektron
değişimi(demirin redoks aktivitesi) bir taraftan gerekli ve yararlı
olurken, diğer taraftan demir fazlalığı durumlarında oluşan serbest
demirin prooksidan özellik göstermesiyle, serbest oksijen radikallerinin
oluşumuna neden olmaktadır. Antioksidanlar tarafından yeterince
detoksifiye edilemeyen serbest oksijen radikalleri, özellikle de hidroksil
radikali, hücresel elemanlar için oldukça zararlı ve toksiktir. (Fenton ve
Heber-Weis reaksiyonları). Bunu önlemek için, vücutta demir hiçbir
zaman serbest bırakılmamaya çalışılır.
Demir İçeren Gıdalar
Proteinden zengin besinler özellikle sırasıyla sakatatlar, dana eti, koyun
eti ve tavuk eti hem demirden zengin besinlerdir hem de içerdikleri
demir emilimiyüksektir. Etlerden başka iyi pişmiş olan kurubaklagiller,
3.
soya fasulyesi, yumurta,kuru meyveler ( özellikle kuru üzüm, kuru
kayısı), pekmez, yeşil sebzeler (ıspanak ), fındık, fıstık, susam, tahin gibi
yiyecekler demirden zengindir.
• Karaciğer gibi sakatatlardaki demirin %30’ u ,
• Dana ve koyun etindeki demirin % 25’ i,
• Tavuk ve balıktaki demirin % 20’ si,
• Yumurtadaki demirin % 15’i,
• Kurubaklagillerdeki demirin % 20’ si,
• Yeşil sebzelerdeki demirin % 8’ i,
• Tahıllardaki demirin % 4’ü emilir.
Demirin Emilimi
Demir, duodenumdan(dahabüyük oranda)ve proksimal jejunumdan
emilir. Musin; demir molekülünü bağlayabilme yeteneği olan bir
proteindir. Musin-demir kompleksi, duodenumun alkali ortamında
emilim için gereklidir.Böylece demir ince bağırsakhücrelerin yüzeyinde
bulunan “integrinler”aracılığıile membrandan geçer ve sitoplazmik bir
demir bağlayıcıprotein olan “mobilferrin” ile bağlanır. Her bir molekül
mobilferrin, bir molekül demir bağlar. Mobilferrin, demiri hücrenin
bazolateral yüzeyine taşır.
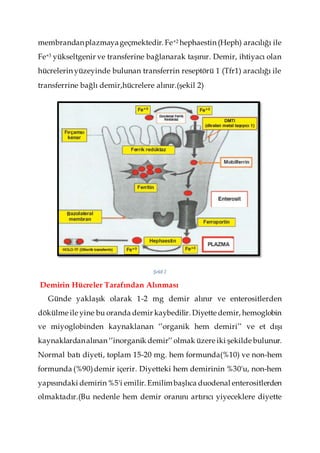
Diyetle alınan Fe+3, duodenumun fırçamsı kenarında duodenal ferrik
redüktaz (Dcytb) ile Fe+2 ye indirgenir ve divalent metal transporter 1
(DMT1) aracılığı ile fırçamsı kenar membranından enterositlere
alınmaktadır. Fe+2 hücrede ferritin olarak depolanıp, dökülen
enterositlerle birlikte atılır ya da ferroportin aracılığı ile basolateral
4.
membrandanplazmayageçmektedir. Fe+2 hephaestin(Heph) aracılığı ile
Fe+3 yükseltgenir ve transferine bağlanarak taşınır. Demir, ihtiyacı olan
hücrelerin yüzeyinde bulunan transferrin reseptörü 1 (Tfr1) aracılığı ile
transferrine bağlı demir,hücrelere alınır.(şekil 2)
Şekil 2
Demirin Hücreler Tarafından Alınması
Günde yaklaşık olarak 1-2 mg demir alınır ve enterositlerden
dökülme ile yine bu oranda demir kaybedilir. Diyette demir, hemoglobin
ve miyoglobinden kaynaklanan ‘’organik hem demiri’’ ve et dışı
kaynaklardanalınan ‘’inorganik demir’’olmak üzere iki şekilde bulunur.
Normal batı diyeti, toplam 15-20 mg. hem formunda(%10) ve non-hem
formunda (%90)demir içerir. Diyetteki hem demirinin %30'u, non-hem
yapısındaki demirin %5'i emilir. Emilimbaşlıca duodenal enterositlerden
olmaktadır.(Bu nedenle hem demir oranını artırıcı yiyeceklere diyette
5.
daha çok yerverilmesi demir eksikliğini önlemede önemli bir yer
tutacaktır.)
Et yemekle alınan hem demiri ve et dışı kaynaklardan alınan
inorganik demirin emilim yolları birbirinden tamamen farklıdır:
1.Non-hem Demirin Emilimi
Non-hem demirin yapısındaki demir, ferrik(+3)yapılarhalindedir.Bu
formu biyolojik olmadığı için, Fe3+ intestinal epitelden transport
olmadan önce, koenzim olarak C vitamini kullanan ferritin redüktaz
enzimi tarafından Fe2+ formuna indirgenmelidir. Bu işlem mide asidi
varlığındagerçekleştirilir. İndirgenme reaksiyonusonunda çinko, bakır,
kobalt gibi metal iyonlarını da taşıyan divalan metal transporter 1
(DMT1) tarafından enterosite demir alımı gerçekleştirilir. DMT1; non-
hem demir alımını sağlayan en önemli proteindir. (DMT 1 ve ferritin
redüktaz sentezi, demir eksikliğinde artmaktadır.) Vitamin C, fruktoz,
sitrat ve aminoasitler ile non-hem demirin absorbsiyonu artar.
Tetrasiklin,proton pompa inhibitörü, fitat(yüksek lifli diyet), kalsiyum,
fosfat, tannat, oksalat ve fenolik bileşikler(çay, kahve) ise non-hem
demirinin absorbsiyonunu engeller. H. pylori enfeksiyonunun yaptığı
gastrik atrofi de, demir eksikliği anemisiyle sonuçlanabilmektedir.
Enterosite alınan demirin bir kısmı ferritin şeklinde depolanır ve
duodenal eksfoliasyon ile atılır. Organizmaya demir alınacaksa,
gereksinim varsa,absorbe edildikten sonra enterositin bazolateral
tarafına taşınır ve oradan insanda bilinen tek demir atıcısı olan
ferroportin ile plazmadaki transferine yüklenir. Fakat önce
seruloplazmin homoloğu ve bir transmembran proteinini olan
hephaestin ile FeII, FeIII haline okside edilmelidir.
6.
2.Hem Demirinin Emilimi
Ettebulunan hemoglobin, barsakta intestinal enzimlerle hem ve
globuline ayrılmakta, globulin yıkım ürünleri hem ve inorganik demiri
solubul halde tutarak absorbsiyonukolaylaştırmaktadır. Hem demirinin
emilimi için, duodenal düşük pH ve emilimikolaylaştıran askorbik asit,
sitrik asit gibi faktörlere gereksinim yoktur. Besinlerde bulunan demir
bağlayıcılardan da etkilenmez. Sadece kalsiyumun emilimi olumsuz
olarak etkilediği gösterilmiştir.
Hem, duodenal enterosite eskiden ‘’hem vesikülü’’ denilen şimdi ise
yeni tanımlanan ‘’hem taşıyıcı protein 1’’ denilen özel bir taşıyıcı ile
girer. Bir kere enterositiçine girdiğinde, hem oksijenaz tarafından Fe2+
molekülü açığa çıkarılır. Çoğu Fe2+, intestinal epitelyum hücresine
girince ferroportin 1 tarafından basolateral membrana salınmaktadır ve
hephaestin tarafından Fe3+’e çevrildikten sonra plazmada transferine
(Tf) bağlanmaktadır. (Ferroportin ayrıca hepatositler, makrofajlar (RES)
ve plasental sinsityotrofoblastlar (fetal sirkulasyona demir girişinin
kontrolü) tarafından da eksprese edilmektedir.)
Demirin Hücreler Tarafından Alınması
Demir, plazmadakaraciğerde sentezlenen ve glukoprotein yapısında
olan transferin tarafından taşınır. Enterositin bazolateral tarafından
ferroportin ile dışarı verilen ve hephaestin ile okside edilerek ferrik
(FeIII) hale getirtildikten sonra transferine yüklenen demir portal
dolaşımdan, çoğu kemik iliğindeki eritrosit öncü hücrelerine olmak
üzere, hücrelere taşınır. Her transferin molekülü, iki tane Fe III’ü güçlü
bir şekilde bağlar. (Hephaestin eksikliğinde, duedenal enterositlerde
7.
demir fazlalığıve demiremilim bozukluğuna bağlıhipokrom mikrositer
anemi olduğu gösterilmiştir.)
Hücreler, tipine göre demiri farklı yollardan alırlar:
1.Enterositler; diyetdemirini apikal tarafta bulunan DMT1 ile alınırken,
2.Makrofajlar;demirifagosite ettikleridolaşımdaki yaşlı eritrositlerdeki
hemoglobinden alırlar. Eritrosit lizisi ile açığa çıkan hemoglobinden
hemoksijenaz ve biliverdin redüktaz ile demir ve bilirubin oluşur.
Makrofajların vakuolar membranlarından demir transportu gene DMT1
ile olmaktadır. Makrofajlardaaçığa çıkan demir, ya tekrar organizmada
dolaşan demir olması için makrofaj ferroportini ile plazmaya verilir ya
da makrofaj içinde ferritin şeklinde depolanır. Ferroportin enterositte
olduğu gibi hücrenin tek demir atıcısıdır. Makrofajdan demir plazmaya
verilirken transferine yüklenebilmesi için gene FeIII şekline getirilmeli
okside edilmelidir. Bu oksidasyon ve tansferrine yüklenme işinde
plazmada bakıra bağlı ferrioksidaz olan ve karaciğerde sentezlenen
seruloplazmin rol alır.
3.Hepatositlerin demir alımı; TfR1 ve TfR2 ile olur. Hepatositler, portal
dolaşımdan aldıklarıdemiri depolarlar ve gerektiğinde ferroportin yolu
ile tekrar dolaşıma verirler.
8.
Şekil 3
Bunlar dışındatüm hücreler, demiriyüzeylerinde bulunan transferin
reseptörlerini kullanarakplazmatransferrininden almaktadırlar.Normal
şartlarda transferin demirle saturasyonu (TS) % 30 orandadır.
(Transferinin demir bağlamakapasitesi tamamen dolduğunda plazmada
serbest transferine bağlı olmayan demir oluşur. Bu demir özellikle
karaciğer ve kalp hücrelerine kolaylıkla girebilir ve hücresel düzeyde
hasar oluşturabilir.)
Plazmadaki diferrik transferin (holotransferrin), hücrenin
membranında, hücre içi demir ihtiyacına göre miktarı belirlenen
düzeyde sentezlenmiş ve hücre yüzeyine yerleşmiş transferrin
resöptörüne (TfR) bağlanır. Tf-TfR1 kompleksi hücreye alınır ve bir
endozom oluşur. Endozom içi pH düşürülerek, transferrin demirden
ayrılır ve demir redükte edilir. Endozomal membrandan demirin(Fe+2)
sitoplazmaya geçişi DMT1ile olur. Sitoplazmadaki demir mitokondride
hem sentezine katılabilir, ferrritin şeklinde depolanabilir veya diğer
metabolik yollarda kullanılabilir. Demirini bırakmış transferrin yani
9.
apotransferrin-TfR reseptör kompleksitekrar hücre yüzeyine gönderilir
ve transferrin tekrar kullanılmak üzere plazmaya salınır.
Transferin Reseptörü
Transferin reseptörü disülfid bağları ile bağlı iki subünitten
oluşmuştur. Reseptörünün eksterasellüler parçası, serumda bulunur.
(Serum transferrin reseptör (sTfR) düzeyi ölçümü, direkt olarak
organizmanın demire olan ihtiyacınını göstermektedir. Ayrıca serum
TfR’nin kaynağı olgunlaşan eritroid hücrelerden dökülen TfR olduğu
için de serumda sTfR ölçümü; plazma demir döngüsü ve eritropoetik
aktivite düzeyini tahmin etmek amacıyla kullanılabilir.) İki ayrı genle
kodlanan TfR1 ve TfR2 şeklinde, iki farklı TfR’ü vardır:
a.TfR1; enterositlerde kript bazolateral kısımda bulunur ve demiri
transferinden alan tüm hücrelerde, en çok kemik iliği eritrosit
öncülerinde bulunur.
b.TfR2; TFR1’in homoloğu olup diferrik transferine bağlanır. TfR1’ in
tersine tüm hücrelerde bulunmaz.En çok karaciğerde,kan hücrelerinde,
duodenal kript hücrelerinde bulunan TfR2, karaciğere demir depoları
sinyallerini iletmede önemlidir. (TfR2 gen mutasyonunun herediter
hemakromatosise yol açması, hepsidin ile ilişkisi belirlenmiştir.)
Organizmadaki Demir Dengesi
Günlük üretilen 300 milyon adet eritrosit için gerekli demirin (20-30
mg) çoğu, makrofajlardaki demir döngüsünden elde edilir. Bu sebeple
günlük 1-2 mg. demir emilimi ancak günlük demir kaybının
karşılayabileceğinden, eritropoezis için kemik iliğinin demir ihtiyacının
karşılanmasında demirin vücuttaki iç döngüsü çok önem taşır.
10.
Hücresel düzeyde demirdüzenlenmesinin moleküler kontrolü;
demirin taşınması, depolanması, kullanımıile ilgili tüm ana proteinlerin
sentezi posttranskripsional düzeyde hücre içi demirle düzenlenmektedir.
Bu düzenlenme hücresel demir sensör proteinleri olan iron regulatuar
proteinler (IRP; sitoplazmada bulunan ve hücre içinde demiri algılayan)
ile iron responsive elementler (IRE ;demir proteinlerinin mRNA’ları
üzerinde bulunan 30 nükleotidlik bölgeyi içerir.) arasındaki ilişkiye
bağlıdır.
Hücre içinde demir eksikliği olduğunda, IRP’ler ile IRE’ler
bağlanırlar. Bu bağlanma; transferin reseptörü (TfR) ve DMT1’in
degredasyonunu azaltır, translasyonunu artırır, ferritin, ferroportin ve
aminolevulinik asit sentaz (eALA-S)’ın sentezlerini durdurur.
Hücre dışı demir konsantrasyonu normal sınırlarda iken hücresel
demir dengesi, IRP/IRE sistemi ile düzeyleri ayarlanan proteinlerle
düzenlenmektedir.
Hücresel demir fazlalığında ise, IRP yapısal olarak değişip IRE’lere
bağlanamayacağıiçin TfR mRNA stabilizasyonubozulur, degredasyonu
artar ve böylece hücre demir alımı dururken, ferritin sentezi artarak
demir depolanır.
Hepsidin ve Organizmanın Demir Dengesi
Hepsidin; esas olarak karaciğerden sentezlenen antimikrobial özellik
gösteren(Hepsidin/ ferroportin sistemi patojenlerin demiri almalarını
engelleyerek konakçı savunmasına katkı sağlar.), dolaşımda bulunan,
idrarla atılan, sistemik demir dengesinin ana düzenleyicisi rolünde 25
11.
aminoasitlikbir hormondur. 19.kromozomda bulunan HAMP geninde
kodlanır. (HAMP geni mutasyonu ile ağır demir birikimi olur.)
Hepsidin sentezi, demir tarafından transkripsiyonel olarak
düzenlenir. Bu düzenlemeyi; kemikteki morfojenik proteinler olan (
BMDs)- SMAD ile ve STAT (signal transducer and activator of
transcription) proteinleri aracılığıyla enflamatuar sitokinlerle yapar.
Hepsidin demir metabolizmasındaki düzenlemeyi, demirin
kullanımını ve depolanmasını koordine ederek ve demirin plazmaya
çıkışını engelleyerek yapmaktadır. Depo demirine ve eritropoezisin
ihtiyacına göre, demiri hücreden dışarı çıkaran hücre yüzeyindeki
ferroportin ekspresyonunu kontrol eder.
Hepsidinin reseptörü, bir bazolateral transmembran proteini olan
ferroportindir.Ferroportin demirin hücreden plazmaya çıkarılmasını ve
hephaestinin yardımı ile plazma transferinine yüklenerek taşınmasını
sağlar.Hepsidinin ferroportine bağlanması, onun internalizasyonuna ve
lizozomal degredasyonuna yol açar. Bunun sonucunda, ferroportinin
membrandan kaybına yol açar.
Anemi ve hipokside; hepsidin sentezi azalır, hücre yüzeyinde
ferroportin artar. Bunun sonucunda demir emilimi ve makrofajlardan
dolaşıma tekrar verilen demir miktarı artar.
Hepsidin/ferroportin sistemi ayrıca patojenlerin demiri almalarını
engelleyerekkonakçısavunmasınakatkısağlar. Enfeksiyon enflamasyon
anemisinde oluşan hipoferrinemi, hastanın savunma
mekanizmalarından biridir. Bu durumdan sorumlu hormon da
hepsidindir. IL-6 ve diğer sitokinlerle hepsidinin arttığı, hemoglobin
12.
sentezi ve eritropoeziçin kullanılacak demiri, demir emilimini
engelleyerek ve retiküloendotelyal sistemde demir blokajı yaparak
azalttığı, hepsidininarttığıbütün durumlardaanemi olduğuçeşitli klinik
durumlarda gösterilmiştir.(şekil 3)
Hepsidininin hipoferrinemi yapıcı etkisi yanında, eritroid öncü
hücrelerinin çoğalmalarınıve yaşamlarınıbozarakeritropoezibaskılayıcı
etkisi de gösterilmiştir.(Enfeksiyonlarda oluşan süperoksit ve hidrojen
peroksitin demir regülatuar protein’in (IRP), demir responsive
element’lere (IRE) bağlanmasını azaltarak, demir metabolizmasına
olumsuz etkileri de bilinmektedir.)
Şekil 4
Demir Metabolizması Bozuklukları
Kalıtsal demir metabolizması bozuklukları, demirin emiliminden
protoporfirin IX’a demir insersiyosununa kadar birçok demir
13.
metabolizması basamağında olabilmeklebirlikte sıklıkla hepsidin
yetersizliğine veya hepsidin artışına bağlı ortaya çıkar.
1.Hemakromatozis:Hemokromatozisin moleküler patogenezi, hepsidin
eksikliği ya da hepsidin rezistansıyla ilişkilidir.
2.Dominant Ferroportin Hastalığı: Heterojen bir prezentasyonu vardır,
iki tipi vardır. Mutantferroportinin plazmamembranına ulaşmadığında
(tip A), demir makrofajlarda birikir ve eritropoez demir demirden
alıkoyulmuş şekilde olabilir. Eğer mutasyon hepsidin ayrışmasında bir
protein rezistansıgeliştirirse(tipB), demir aynıhemokromatozisteki gibi
birikir.
3.Demire Refrakter Demir Eksikliği Anemisi(IRIDA):Aşırı hepsidin
üretimine bağlı bir hastalıktır. IRIDA, karaciğerden sentezlenen tip 2
transmembranserin proteaz olan matriptase-2 yi kodlayan TMPRSS6’nın
mutasyonuna bağlı resesif bir bozukluktur. Genom bazlı yapılan
çalışmalar, TMPRSS6 ‘nın, serum demiri, transferrin saturasyonu,
hemoglobin ve eritrosit varyasyonlarındaki genetik kontrol üzerinde
rolü olduğunu göstermiştir.
3.Hipotransferrinemi:Hipotransferrinemi saptanamaz düzeyde ya da çok
düşük transferrinle ve demir eksikliği ve hemokromatozis bulgularıyla
karakterize ender resesif bir hastalıktır.
4.İki değerlikli metal taşıyıcı 1 mutasyonları (DMT1):DMT1 hastalığı
DMT1 genindeki mutasyona bağlı olan otozomal dominant ender bir
hastalıktır.
14.
5.Sideroblastik anemiler:Sideroblastik anemilereritroblastların
perinükleer mitokondrilerinde demir birikir.
6.Aserüloplazminemi:Aserüloplazminemi ilk olarak Japonya’da ortaya
çıkmış ve daha sonra sporadik olarak tüm dünyada görülmüş resesif bir
hastalıktır. Hastalık, demirve bakır metabolizmasını birbirine bağlayan
ve bakır içeren bir plazma proteini olan seruloplazmin (Cp)
mutasyonuna bağlıgelişir.imine bağlı ring sideroblastlarla karakterize
heterojen bir hastalıktır.
7.İzole Hiperferritinemi:Ailesel hiperferritineminin nadir formları,
yüksek ferritin seviyeleri, anemisiz normal transferin saturasyonu,
yüksek demir düzeyleri ve klinik semptomlarla seyreder.
8.Hiperferritenemi-Katarakt Sendromu:IRE-IRP sisteminin
deregülasyonuna örnektir.
9.Benign Hiperferritinemi:Nadir görülürler, aileseldir, katarakt
görülmez. L-ferritin molekülünü sekresyona daha duyarlı hale getiren
Thr30I1e’mutasyonu sonucu meydana gelir.
10.Nöroferritinopati:L-ferritinin C terminalindeki nukleotid
insersiyonları dominant bir nadir hastalık olan nörodegerenasyona ve
nöroferritinopatiye neden olur. Sebebi oksidasyon artışı ve anormal
protein agregasyonuna bağlı hücre ölümüdür.
Demir metabolizması ile ilgili mitokondriyal bozukluklar
Mitokondriyal demir metabolizması ile ilgili güncel bilgiler, bazı
hastalıklardaki patolojik mekanizmaları açıklamak için yol gösterir. Bu
15.
hastalıklar genellikle eritrositserisi etkilediği gibi nöral dokuyu da
etkiler.
1.Friedreich Ataksisi:Hastalarında yapılan kalp biyopsilerinde, demir
sülfür kümeleri içeren proteinlerin azalması( akonitaz ve mitokondriyal
respiratuar zincir kompleksi) bu bulgunun aşırı demir yüklenmesine
bağlı mitokondriyal hasar ile ilişkili olduğunu göstermiştir.
2.X’e Bağlı Sideroblastik Anemi:X’e bağlı sideroblastik anemi, ALAS2
geninde görülen mutasyona bağlı nadir görülen bir hastalıktır.
3.GLRX5 Mutasyonuna Bağlı Pridoksine Duyarsız Sideroblastik
Anemi:GLRX5 eksikliği demir sülfür kümelerinin sentezinin
bozulmasına neden olur, bu da sitoplazmik İRP1 i aktifler ve sonuç
olarak Sellüer demir alımı artar.
4.Ataksi ile Birlikte Olan X’e Bağlı Sideroblastik Anemi:Nükleer
genomdaki mutasyona bağlı mitokondriyal hastalıktır. Mitokondride,
bilinmeyen bir mekanizmayla demir birikimi oluşur.
5.ISCU1 Defektiyle İlişkili Miyopati:Friedreich ataksisindeki ve
pridoksine duyarsız GLRX5 sideroblastik anemisindeki gibi ISCU1
defektiyle ilişkili miyopati de demir sülfür kümeleri eksikliğine neden
olur.
6.Gracile Sendromu:Gracile Sendromu(büyüme geriliği,aminoaasidüri,
kolestaz, demir birikimi, laktik asidoz ve erken ölüm) sıklıkla
Finlandiya’daki yenidoğanlarda bildirilmiştir. Kadavra bulguları
karaciğerde demir fazlalığı, intrahepatiksafra kanallarındaazlık ve daha
büyük hastalarda (4 aylık) periportal fibrozisi göstermektedir.
16.
Kaynaklar
ConradME, UmbreitJN. A concise review: İron absorption-the mucin-mobilferrin-integrin pathway. A
competitive pathway for metal absorption. Am J Hematol 1993;42:67-73.
Fleming RE, Bacon BR. Orchestration of İron Homeostasis. NEngl J Med 2005;352:1741-4.
Ganz T, Nemeth E. Regulation of iron acquisition and iron distribution in mammals. Biochim Biophys
Acta 2006;1763:690-9.
Mackenzie B, Garrick MD. Iron Imports. II. Iron uptake at the apical membrane in the intestine. Am J
Physiol Gastrointest Liver Physiol 2005;289:981-6.
Ma Y, Yeh M, Yeh KY, Glass J. Iron Imports. V. Transport of iron through the intestinal epithelium. Am J
Physiol Gastrointest Liver Physiol 2006;290:417-22.
West AR, Oates PS. Mechanisms of heme iron absorption: current questions and controversies. World J
Gastroenterol 2008;14:4101-10.
Donovan A, Brownlie A, Zhou Y, Shepard J, Pratt SJ, Moynihan J, et al. Positional cloning of zebrafish
ferroportin 1 identifies a conserved vertebrate iron exporter. Nature 2000;403:776-81.
Nemeth E, Preza GC, Jung CL, Kaplan J, Waring AJ, Ganz T. The N-terminus of hepcidin is essential for
its interaction with ferroportin: structure-function study. Blood 2006;107:328-33.