Download free for 30 days
Sign in
Upload
Language (EN)
Support
Business
Mobile
Social Media
Marketing
Technology
Art & Photos
Career
Design
Education
Presentations & Public Speaking
Government & Nonprofit
Healthcare
Internet
Law
Leadership & Management
Automotive
Engineering
Software
Recruiting & HR
Retail
Sales
Services
Science
Small Business & Entrepreneurship
Food
Environment
Economy & Finance
Data & Analytics
Investor Relations
Sports
Spiritual
News & Politics
Travel
Self Improvement
Real Estate
Entertainment & Humor
Health & Medicine
Devices & Hardware
Lifestyle
Change Language
Language
English
Español
Português
Français
Deutsche
Cancel
Save
Submit search
EN
Uploaded by
Garsiet Creus
2,998 views
Biology Chapter10
Education
◦
Read more
3
Save
Share
Embed
Embed presentation
Download
Downloaded 34 times
1
/ 70
2
/ 70
Most read
3
/ 70
Most read
4
/ 70
5
/ 70
6
/ 70
7
/ 70
8
/ 70
9
/ 70
10
/ 70
11
/ 70
12
/ 70
13
/ 70
14
/ 70
15
/ 70
16
/ 70
17
/ 70
18
/ 70
19
/ 70
20
/ 70
21
/ 70
22
/ 70
23
/ 70
24
/ 70
25
/ 70
26
/ 70
27
/ 70
28
/ 70
29
/ 70
30
/ 70
31
/ 70
32
/ 70
33
/ 70
34
/ 70
35
/ 70
36
/ 70
37
/ 70
38
/ 70
39
/ 70
40
/ 70
41
/ 70
42
/ 70
43
/ 70
44
/ 70
45
/ 70
46
/ 70
47
/ 70
48
/ 70
49
/ 70
50
/ 70
51
/ 70
52
/ 70
53
/ 70
54
/ 70
55
/ 70
56
/ 70
57
/ 70
58
/ 70
59
/ 70
60
/ 70
61
/ 70
62
/ 70
63
/ 70
64
/ 70
65
/ 70
66
/ 70
67
/ 70
68
/ 70
69
/ 70
70
/ 70
More Related Content
PDF
ระบบหายใจ
by
Thitaree Samphao
PDF
ย่อยอาหาร
by
Wichai Likitponrak
PDF
16แบบทดสอบความหลากหลายทางชีวภาพ
by
สำเร็จ นางสีคุณ
PDF
ลำต้น54
by
Oui Nuchanart
DOCX
ใบงานคลื่นกล ม.5 .docx
by
sathanpromda
PDF
เฉลยชีววิทยาหน้า52- 59
by
Wan Ngamwongwan
PDF
ระบบย่อยอาหาร และ การสลายอาหารเพื่อให้ได้พลังงาน
by
พัน พัน
PDF
แบบทดสอบดาราศาสตร์ ม.3
by
Jariya Jaiyot
ระบบหายใจ
by
Thitaree Samphao
ย่อยอาหาร
by
Wichai Likitponrak
16แบบทดสอบความหลากหลายทางชีวภาพ
by
สำเร็จ นางสีคุณ
ลำต้น54
by
Oui Nuchanart
ใบงานคลื่นกล ม.5 .docx
by
sathanpromda
เฉลยชีววิทยาหน้า52- 59
by
Wan Ngamwongwan
ระบบย่อยอาหาร และ การสลายอาหารเพื่อให้ได้พลังงาน
by
พัน พัน
แบบทดสอบดาราศาสตร์ ม.3
by
Jariya Jaiyot
What's hot
PDF
โครงงานเเยมกล้วย(Complete)
by
Pongpan Pairojana
PDF
โครงงานวิทยาศาสตร์ ถ่านอัดแท่งสมุนไพร1
by
Chok Ke
PDF
ใบงานที่ 1 ธาตุและสารประกอบ
by
website22556
PDF
ใบงานวิทยาศาสตร์ ป.5 ครบ
by
ssuserf8d051
PDF
ข้อสอบโครงงาน ม 2
by
Weerachat Martluplao
PDF
แบบฝึกหัด
by
Hansa Srikrachang
DOCX
ใบงานที่ 4.1 ม4
by
mrtv3mrtv4
PDF
โครงงาน 5 บท
by
ศุภกรณ์ วัฒนศรี
PDF
เซลล์หน่วยของสิ่งมีชีวิต
by
Popeye Kotchakorn
PDF
ระบบย่อยอาหาร - Digestive system
by
supreechafkk
PPTX
กฎหมายทะเบียนราษฎร
by
Yosiri
DOC
แบบฝึกหัดที่ 2 เซลล์พืช และเซลล์สัตว์
by
Wann Rattiya
PDF
ใบงานที่ 13 การแบ่งเซลล์แบบไมโทซิส
by
Aomiko Wipaporn
PDF
1.ตัวอย่างแผนบูรณาการเศรษฐกิจพอเพียง
by
Wareerut Hunter
PPTX
สิ่งมีชีวิตกับสิ่งแวดล้อม ประถม
by
Ta Lattapol
PDF
การปฏิสนธิและการเจริญเติบโตของพืชดอก
by
Thanyamon Chat.
PDF
Lesson3 plantgrowth2
by
Wichai Likitponrak
PDF
พืชใบเลี้ยงเดี่ยวและพืชใบเลี้ยงคู่
by
Benjapron Seesukong
PDF
ห้องสีขาว 1.1
by
peter dontoom
PDF
12 ไอเดียสอนเด็กเรื่องอาชีพให้สนุก
by
Influencer TH
โครงงานเเยมกล้วย(Complete)
by
Pongpan Pairojana
โครงงานวิทยาศาสตร์ ถ่านอัดแท่งสมุนไพร1
by
Chok Ke
ใบงานที่ 1 ธาตุและสารประกอบ
by
website22556
ใบงานวิทยาศาสตร์ ป.5 ครบ
by
ssuserf8d051
ข้อสอบโครงงาน ม 2
by
Weerachat Martluplao
แบบฝึกหัด
by
Hansa Srikrachang
ใบงานที่ 4.1 ม4
by
mrtv3mrtv4
โครงงาน 5 บท
by
ศุภกรณ์ วัฒนศรี
เซลล์หน่วยของสิ่งมีชีวิต
by
Popeye Kotchakorn
ระบบย่อยอาหาร - Digestive system
by
supreechafkk
กฎหมายทะเบียนราษฎร
by
Yosiri
แบบฝึกหัดที่ 2 เซลล์พืช และเซลล์สัตว์
by
Wann Rattiya
ใบงานที่ 13 การแบ่งเซลล์แบบไมโทซิส
by
Aomiko Wipaporn
1.ตัวอย่างแผนบูรณาการเศรษฐกิจพอเพียง
by
Wareerut Hunter
สิ่งมีชีวิตกับสิ่งแวดล้อม ประถม
by
Ta Lattapol
การปฏิสนธิและการเจริญเติบโตของพืชดอก
by
Thanyamon Chat.
Lesson3 plantgrowth2
by
Wichai Likitponrak
พืชใบเลี้ยงเดี่ยวและพืชใบเลี้ยงคู่
by
Benjapron Seesukong
ห้องสีขาว 1.1
by
peter dontoom
12 ไอเดียสอนเด็กเรื่องอาชีพให้สนุก
by
Influencer TH
Similar to Biology Chapter10
PDF
การลำเลียงน้ำของพืช
by
Anana Anana
PDF
การลำเลียงอาหารของพืช
by
Anana Anana
PDF
การลำเลียงสารอาหารของพืช
by
Anana Anana
PDF
ธนาคารความรู้ การดำรงชีวิตของพืช
by
Kunnanatya Pare
PPT
การลำเลียงน้ำของพืช
by
nokbiology
PPT
การลำเลียงน้ำของพืช
by
nokbiology
PDF
การคายน้ำของพืช
by
dnavaroj
PDF
การลำเลียงน้ำและอาหารของพืช
by
Thanyamon Chat.
PDF
Handling and nama plant
by
Miss.Yupawan Triratwitcha
PDF
ขนราก
by
dnavaroj
PDF
ระบบลำเลียงน้ำและแร่ธาตุในพืช
by
dnavaroj
PPTX
12.การคายน้ำและการแลกแก๊สลำเลียงพืช
by
Wichai Likitponrak
PDF
การลำเลียงอาหารในพืช
by
dnavaroj
PPT
การลำเลียงน้ำของพืช
by
Nokko Bio
PPT
การลำเลียงธาตุอาหารของพืช
by
Nokko Bio
PPT
Responseของพืช
by
Issara Mo
PDF
การแลกเปลี่ยนแก๊สและการคายน้ำ
by
Anana Anana
PPT
การลำเลียงน้ำของพืช
by
Nokko Bio
PDF
หน่วยการเรียนรู้ที่-10-การลำเลียงของพืช.pdf
by
NoeyWipa
PDF
สรุปเซลล์
by
สายฝน ต๊ะวันนา
การลำเลียงน้ำของพืช
by
Anana Anana
การลำเลียงอาหารของพืช
by
Anana Anana
การลำเลียงสารอาหารของพืช
by
Anana Anana
ธนาคารความรู้ การดำรงชีวิตของพืช
by
Kunnanatya Pare
การลำเลียงน้ำของพืช
by
nokbiology
การลำเลียงน้ำของพืช
by
nokbiology
การคายน้ำของพืช
by
dnavaroj
การลำเลียงน้ำและอาหารของพืช
by
Thanyamon Chat.
Handling and nama plant
by
Miss.Yupawan Triratwitcha
ขนราก
by
dnavaroj
ระบบลำเลียงน้ำและแร่ธาตุในพืช
by
dnavaroj
12.การคายน้ำและการแลกแก๊สลำเลียงพืช
by
Wichai Likitponrak
การลำเลียงอาหารในพืช
by
dnavaroj
การลำเลียงน้ำของพืช
by
Nokko Bio
การลำเลียงธาตุอาหารของพืช
by
Nokko Bio
Responseของพืช
by
Issara Mo
การแลกเปลี่ยนแก๊สและการคายน้ำ
by
Anana Anana
การลำเลียงน้ำของพืช
by
Nokko Bio
หน่วยการเรียนรู้ที่-10-การลำเลียงของพืช.pdf
by
NoeyWipa
สรุปเซลล์
by
สายฝน ต๊ะวันนา
More from Garsiet Creus
PDF
โครงการเรียนล่วงหน้า รุ่นที่ 10 เพื่อเข้าศึกษาในสังกัดคณะของมหาวิทยาลัยเกษตร...
by
Garsiet Creus
DOCX
สารชีวโมเลกุล By Prof.Dr.Garsiet Creus
by
Garsiet Creus
PDF
วศ.บ.(วิศวกรรมหุ่นยนต์และระบบอัตโนมัติ) ฟีโบ้
by
Garsiet Creus
PDF
แนวข้อสอบความรู้ความสามารถทั่วไป 30 ข้อ
by
Garsiet Creus
DOC
ข้อสอบความรู้รอบตัว
by
Garsiet Creus
DOC
ข้อสอบพร้อมเฉลยข้อสอบภาค.ก. ภาษาไทย
by
Garsiet Creus
PDF
แนวข้อสอบความเข้าใจ อุปมา อุปมัย 62 ข้อ
by
Garsiet Creus
PDF
แนวข้อสอบการอ่านคำ 106 ข้อ
by
Garsiet Creus
PDF
แนวข้อสอบการรู้จักความหมายของคำและกลุ่มคำ 30 ข้อ
by
Garsiet Creus
PDF
แนวข้อสอบการใช้คำศัพท์และหลักของภาษาไทย 198 ข้อ
by
Garsiet Creus
DOC
ข้อสอบพร้อมเฉลยข้อสอบภาค.ก.
by
Garsiet Creus
DOC
เทคนิคการทำข้อสอบ ก.พ.
by
Garsiet Creus
PDF
B.eng. (civil engineering) inter
by
Garsiet Creus
PDF
TEST Aptitude2
by
Garsiet Creus
PDF
TEST Aptitude
by
Garsiet Creus
DOC
ช่างเขียนแบบ
by
Garsiet Creus
DOC
สรุปสูตรฟิสิกส์
by
Garsiet Creus
DOC
Ch9 wave exercises
by
Garsiet Creus
DOC
Wave
by
Garsiet Creus
PDF
Chapter2
by
Garsiet Creus
โครงการเรียนล่วงหน้า รุ่นที่ 10 เพื่อเข้าศึกษาในสังกัดคณะของมหาวิทยาลัยเกษตร...
by
Garsiet Creus
สารชีวโมเลกุล By Prof.Dr.Garsiet Creus
by
Garsiet Creus
วศ.บ.(วิศวกรรมหุ่นยนต์และระบบอัตโนมัติ) ฟีโบ้
by
Garsiet Creus
แนวข้อสอบความรู้ความสามารถทั่วไป 30 ข้อ
by
Garsiet Creus
ข้อสอบความรู้รอบตัว
by
Garsiet Creus
ข้อสอบพร้อมเฉลยข้อสอบภาค.ก. ภาษาไทย
by
Garsiet Creus
แนวข้อสอบความเข้าใจ อุปมา อุปมัย 62 ข้อ
by
Garsiet Creus
แนวข้อสอบการอ่านคำ 106 ข้อ
by
Garsiet Creus
แนวข้อสอบการรู้จักความหมายของคำและกลุ่มคำ 30 ข้อ
by
Garsiet Creus
แนวข้อสอบการใช้คำศัพท์และหลักของภาษาไทย 198 ข้อ
by
Garsiet Creus
ข้อสอบพร้อมเฉลยข้อสอบภาค.ก.
by
Garsiet Creus
เทคนิคการทำข้อสอบ ก.พ.
by
Garsiet Creus
B.eng. (civil engineering) inter
by
Garsiet Creus
TEST Aptitude2
by
Garsiet Creus
TEST Aptitude
by
Garsiet Creus
ช่างเขียนแบบ
by
Garsiet Creus
สรุปสูตรฟิสิกส์
by
Garsiet Creus
Ch9 wave exercises
by
Garsiet Creus
Wave
by
Garsiet Creus
Chapter2
by
Garsiet Creus
Biology Chapter10
1.
บทที่ 10 การดํารงชีวิตของพืช Maintenance of
Plant Life วัตถุประสงค์การเรียนรู้ เมือศึกษาบทเรียนนี้แล้วนักศึกษาสามารถ ่ 1. อธิบายกลไกการลําเลียงนํ้า แร่ธาตุ และสารอาหารของพืช 2. อธิบายกลไกการสังเคราะห์ดวยแสงของพืช ้ 3. บอกโครงสร้างของดอกและชนิดของดอก 4. อธิบายกระบวนการสร้างเซลล์สบพันธุของพืช ื ์ 5. อธิบายการถ่ายละอองเรณูและการผสมพันธุของพืช ์ 6. บอกส่วนประกอบและชนิดของผล 7. อธิบายการงอกและการเจริญเติบโตของพืช 8. อธิบายกลไกการตอบสนองของพืช 9. อธิบายการทํางานของสารเคมีต่อการตอบสนองของพืช การดํารงชีวตของพืชมีก ระบวนการต่า ง ๆ ได้แ ก่ การลํา เลีย งสาร พืช ลํา เลีย ง ิ สารอนินทรียพวกนํ้าและแร่ธาตุจากดินโดยระบบท่อนํ้า (Xylem) แก๊สคาร์บอนไดออกไซด์ ์ เข้า ทางปากใบ สารอนิ น ทรีย์เ หล่ า นี้ พ ืช นํ า ไปใช้ใ นกระบวนการสัง เคราะห์ ด้ว ยแสง (Photosysthesis) โดยการรวมกันทางเคมีของคาร์บอนไดออกไซด์และนํ้ามีพลังงานแสง เป็ นตัวช่วย ได้เป็ นคาร์โบไฮเดรต และผลพลอยได้ คือ ออกซิเจนและนํ้า สารอาหารทีพช ่ ื สร้างได้จะลําเลียงไปทางท่ออาหาร (Phloem) ไปยังส่วนต่าง ๆ ของพืช กระบวนการเมแทบอลิซม พืชมีการสลายโมเลกุลของสารอาหารคาร์โบไฮเดรต ึ โดยการสันดาปด้วยออกซิเจน เพื่อนํ าพลังงานไปใช้ในกิจกรรมดําเนินชีวตเช่นเดียวกับ ิ สัต ว์ กระบวนการขับ ถ่ า ยของเสีย พืช มีก ารขับ ถ่ า ยของเสีย ในรูป แก๊ ส และไอนํ้ า เข้า สู่ บรรยากาศทางปากใบเป็ นส่วนใหญ่ พืชสามารถเคลื่อนไหวเพือตอบสนองต่อสิงเร้าได้ ทัง ่ ่ ้ สิงเร้าภายนอก เช่น แสงสว่าง และสิงเร้าภายใน คือ ฮอร์โมน พืชมีกระบวนการสืบพันธุ์ ่ ่ CU 474 413
2.
ทังอาศัยเพศ และไม่อาศัยเพศ กระบวนการเจริญเติบโตอาศัย
เนื้อเยื่อเจริญทํา หน้ า ที่ ้ แบ่งตัวเพิมจํานวน และเปลียนแปลงไปเป็ นเนื้อเยือถาวร เพือเพิมขนาดของลําต้น ่ ่ ่ ่ ่ 10.1 การลําเลียงในพืช การลําเลียงนํ้าของพืช (Water Conduction) บริเวณทีมการดูดนํ้ามากทีสด คือ บริเวณขนราก (Root Hair Zone) ซึงมีเซลล์ ่ ี ุ่ ่ ขนรากทําหน้าทีสําคัญ ในดินนํ้ าจะเคลื่อนทีมาที่ผวรากโดยกระบวนการ Mass Flow ่ ่ ิ ซึงเป็ นการเคลื่อนทีของนํ้ าทีถูกผลักดันไปโดยความแตกต่างของความดันนํ้ าสองบริเวณ ่ ่ ่ คือ นํ้าบริเวณใกล้รากจะแพร่เข้าสูเซลล์ของรากตลอดเวลา ทําให้ปริมาณนํ้าบริเวณนี้ลดลง ่ ความดันของนํ้ าจึงลดลงตํ่ากว่าความดันของนํ้ าทีอยู่ห่างจากราก นํ้ าบริเวณนี้จงไหลมาที่ ่ ึ ผิวรากได้ โดยผ่านมาตามช่องว่างระหว่างอนุภาคดินทีตดต่อกัน ่ ิ นํ้ าในดินทีสมผัสกับผิวรากจะเข้าสู่ท่อนํ้ าในรากโดยผ่านเซลล์ชนต่าง ๆ คือ เอพิเดอร์ ่ ั ั้ มิส คอร์เทกซ์ เอนโดเดอร์มส และท่อนํ้า ด้วย 2 วิถทาง ิ ี 1. วิถอะโปพลาสต์ (Apoplast Pathway) นํ้าจะเคลื่อนทีจากเซลล์หนึ่งไปยังอีก ี ่ เซลล์ โดยทาง Membrane หรือทางช่องว่างระหว่างเซลล์ ซึงนํ้าจะต้องเคลื่อนทีขามผ่าน ่ ่ ้ Membrane ในแต่ละเซลล์ 2 ครัง และขณะที่น้ํ าอยู่ในเซลล์น้ํ าอาจจะเข้าและออกจาก ้ แวคิวโอลได้ดวย ้ 2. วิถซมพลาสต์ (Symplast Pathway) นํ้าเคลื่อนทีจากเซลล์หนึ่งไปอีกเซลล์ทอยู่ ี ิ ่ ่ี ข้างเดียว โดยผ่านทางรูผนังเซลล์ (Plastmodesma) ช่องทางนี้เป็ นทางติดต่อของไซโต พลาสซึม ดังนัน นํ้าจึงเคลื่อนทีผานไซโตพลาสซึมจากเซลล์หนึ่งยังอีกเซลล์ วิถซมพลาสต์ ้ ่ ่ ี ิ นํ้าสามารถผ่านเซลล์เอพิเดอร์มสเข้าสูคอร์เทกซ์ เอนโดเดอร์มส และไซเลมได้ ิ ่ ิ การลําเลียงนํ้ าโดยวิถีอะโปพลาสต์ จะมาสิ้นสุดที่เซลล์เอนโดเดอร์มสเนื่องจาก ิ เซลล์ชนนี้มแถบไขมันพวกซูเบอริน (Suberin) เคลือบผนังเซลล์ทเรียกว่า แถบแคสปาั้ ี ่ี เรียน (Casparian Stripe) ซึงไขมันไม่ละลายในนํ้า ทําให้น้ําผ่าน Membrane ไม่ได้ ดังนัน ่ ้ นํ้ า ที่ลํา เลีย งมาโดยวิถีอ ะโปพลาสต์จ ะเข้า สู่เ อนโดเดอร์ม ิส ได้ด้ว ยวิถีซิม พลาสต์ หรือ ช่องว่างของแถบแคสปาเรียนทีอาจเกิดขึนระหว่างการเกิดรากแขนง ่ ้ อย่างไรก็ตาม พบว่า สารซูเบอรินจะมีแต่เฉพาะในเอนโดเดอร์มสของรากทีมอายุ ิ ่ ี มาก ซึงหยุดการเจริญเติบโตแล้ว และเป็ นบริเวณทีห่างจากปลายราก ซึงมีการดูดนํ้าน้อย ่ ่ ่ 414 CU 474
3.
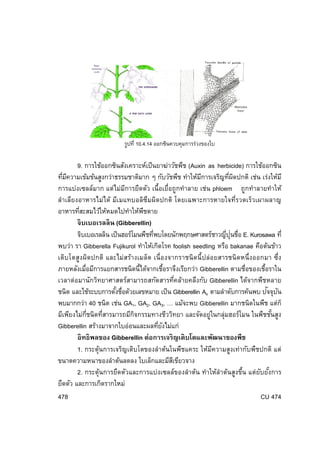
ส่วนปลายรากที่อายุยงน้อยไม่พบซูเบอริน ดังนัน นํ้
าจึงเคลื่อนทีผ่านเซลล์บริเวณปลาย ั ้ ่ รากในปริมาณทีมาก ่ รูปที่ 10.1.1 วิถทางการลําเลียงนํ้าทางรากพืชสองเส้นทาง คือ อะโปพลาสต์และซิมพลาสต์ ี (จาก George B. Johnson, 1997) กระบวนการดูดนํ้าของพืช พืชดูดนํ้าเข้าสูเซลล์ดวย 2 วิธี ได้แก่ ่ ้ 1. กระบวนการออสโมซิส (Osmosis) เป็ นวิธทพชดูดนํ้าได้เป็ นส่วนใหญ่โดยเริม ี ่ี ื ่ จากเซลล์ขนราก ซึงมีสารละลายอยู่มากในไซโตพลาสซึมจึงมีแรงดันออสโมซิส (Osmotic ่ Pressure) สูงกว่านํ้าในดิน นํ้าจากดินจึงแพร่เข้าสูเซลล์ได้ เมือนํ้าเข้าสูเซลล์ ขนรากจะทํา ่ ่ ่ ให้ความเข้มข้นของสารละลายในเซลล์น้อยกว่าเซลล์ทอยู่ถดเข้าไป นํ้าจากเซลล์ขนรากจึง ่ี ั แพร่เข้าสูเซลล์ใกล้เคียงได้ และในทํานองเดียวกันนี้น้ําจะสามารถแพร่ต่อไปยังเซลล์ถดไป ่ ั ได้เรือย ๆ ่ 2. กระบวนการแอกทีฟ แอพซอพชัน (Active Absorption) เป็ นกระบวนการดูดนํ้า ที่พชใช้พลังงานซึ่งได้จากขบวนการเมแทบอลิซึมของเซลล์ดงนํ้ าจากบริเวณที่มความ ื ึ ี เข้มข้นน้อยกว่านํ้าในเซลล์เข้าสูเซลล์ได้ ซึงมักเกิดในบริเวณทีแห้งแล้ง นํ้าในดินมีน้อย ่ ่ ่ กลไกการลําเลียงนํ้าของพืช การลําเลียงนํ้ าของพืชจากส่วนล่าง คือ ราก ขึ้นสู่ท่ีสูงคือ ลํา ต้นและใบของพืช ั ั ่ ปจจุบนเชือว่าพืชมีกลไกการลําเลียงหลายวิธรวมกัน ได้แก่ ี่ 1. แรงดันราก (Root Pressure) คือ แรงดันให้น้ําเคลื่อนทีต่อเนื่องกันจากรากเข้าสู่ ่ ไซเลมจนถึงปลายยอดของพืช แรงดันนี้เป็ นกระบวนการ Active Absorption แรงดันรากมี เพียง 2-5 บรรยากาศเท่านัน และเกิดเฉพาะกับพืชบางชนิดในสภาพพื้นดินที่มค วาม ้ ี CU 474 415
4.
ชุ่มชื้นมาก ดังนันแรงดันรากเพียงอย่างเดียวจึงไม่น่าจะเพียงพอที่จะดันให้น้ํ าขึนไปถึง ้ ้ ยอดทีมความสูงมาก
ๆ ได้ ต้องมีแรงดันอย่างอื่นมาช่วยดันให้น้ําขึนไปด้วย ่ ี ้ 2. คะพิลลารี แอคชัน (Capillary Action) คือ การเคลื่อนทีของนํ้าในหลอดขนาด ่ เล็ก (Capillary Tube) ทีถูกดันให้ขนไปสูงกว่าระดับนํ้าในบริเวณเดียวกัน ซึงเกิดจากแรง ่ ้ึ ่ ตึงผิวของโมเลกุลของนํ้ากับผนังด้านข้างของหลอดทีมแรงยึดเกาะกัน เรียกว่า Adhesion ่ ี คะพิลลารี แอคชัน ใช้อธิบายการเคลื่อนที่ของนํ้ าในท่อเวสเซลที่ไหลต่อกันอย่างไม่ขาด สาย เป็ นเพราะโมเลกุลของนํ้ามีแรงดึงดูดซึงกันและกัน คือ แรงโคฮีชน (Cohesion) และมี ่ ั แรง Adhesion ยึดเกาะกับผิวของท่อเวสเซล นํ้าจะลําเลียงขึนไปได้สงมาก ๆ ท่อจะต้องมี ้ ู ขนาดเล็กมาก แต่ท่อนํ้ าของพืชยังมีขนาดเล็กไม่พอทีจะทําให้เกิดคะพิลลารี แอคชัน นํ า ่ นํ้าไปถึงยอดพืชทีสงมาก ๆ ได้ ู่ 3. ทรานสไปเรชัน พูล (Transpiration Pull) คือ แรงทีเกิดจากการคายนํ้าของพืช ่ ซึงเป็ นวิธลาเลียงนํ้าได้ดของพืชทุกชนิด ทุกขนาด ใบของพืชมีการคายนํ้าตลอดเวลาทําให้ ่ ีํ ี เซลล์ของใบขาดนํ้าจึงเกิดแรงดึงนํานํ้าจากข้างล่างขึนมาทดแทนได้อย่างต่อเนื่อง ้ การคายนํ้าทีใบทําให้เซลล์ทใบขาดนํ้า จึงมีแรงดันออสโมซิสสูงขึน ส่วนแรงดันเต่ง ่ ่ี ้ (Turgor Pressure) จะลดลงเรือย ๆ จนมีค่าเป็ นศูนย์ ใบจะเริมเหียว แต่ยงคายนํ้าได้อก ่ ่ ่ ั ี ถ้าสิงแวดล้อมอํานวยทําให้ค่า Turgor Pressure เป็ นลบ ใบจะเหียวจนเห็นได้ชดเจน ่ ่ ั ขณะเดียวกัน Osmotic Pressure จะยิงเพิมมากขึน จึงมีแรงดัน (Tension) ดึงนํ้าจาก ่ ่ ้ ข้างล่างขึนมาด้านบน เซลล์ดานล่างจะขาดนํ้ าจึงเหียว ทําให้ดูดนํ้าจากเซลล์ล่างถัดไปได้ ้ ้ ่ อีกต่อเนื่องกันไป แสดงว่าการคายนํ้ ามากทําให้มแรงดึงมากขึน นํ้ าจึงขึนสู่ทสงได้ พบว่า ี ้ ้ ่ี ู แรงดึงทีเกิดจากการคายนํ้าของพืชมีมากถึง 200-300 บรรยากาศ ่ การดึงนํ้ าขึนสู่ทสูงย่อมต้องใช้แรงมาก เนื่องจากมีการเสียดทานของเซลล์ต่าง ๆ ้ ่ี แต่พบว่า สายนํ้าไม่ขาดตอนเลย แสดงว่าโมเลกุลของนํ้ามีแรงดึงดูดกัน (Cohesion) มาก ซึงพบว่า มีประมาณ 150 บรรยากาศ ่ แรงทังสามชนิด คือ แรงจากการคายนํ้ า (Transpiration Pull) แรงดึงของนํ้ า ้ (Tention) และแรงยึดเกาะของโมเลกุลของนํ้า (Cohesion) ทําให้น้ําในลําต้นพืชสามารถ ลําเลียงนํ้าจากด้านล่างขึนไปสูยอดสูง ๆ ได้ โดยไม่ขาดสาย ้ ่ Dixon และ Joly ได้ตงทฤษฎี Transpiration Cohesion Tension Theory ในปี ั้ ั ั 1894 ซึงใช้อธิบายการลําเลียงนํ้าของพืช และเชือถือกันมากทีสดจนถึงปจจุบน ่ ่ ุ่ 416 CU 474
5.
การลําเลียงธาตุอาหารของพืช (Translocation of
Minerals) พืชส่วนใหญ่ ดูดเกลือแร่ในรูปของไอออน และเลือกดูดไอออนชนิดต่าง ๆ ด้วย อัตราเร็วไม่เท่ากัน และมีการแข่งขัน วิธการดูดเกลือแร่ของพืชทําได้หลายวิธี ี 1. การแพร่ (Diffusion) ปกติในดินจะมีเกลือแร่มากกว่าในเซลล์เสมอ ดังนัน เกลือ ้ แร่จากดินจึงแพร่เข้าสูเซลล์ได้ เมื่อเซลล์หนึ่งรับเกลือแร่เข้ามาทําให้มความเข้มข้นสูงกว่า ่ ี เซลล์ถดไป จึงแพร่ผานต่อกันไปได้อย่างต่อเนื่อง ั ่ 2. การแลกเปลียนอิออน (Ion Exchange) เป็ นวิธดดเกลือแร่โดยมีการแลกเปลียน ่ ี ู ่ + ่ ไอออนกับผิวราก ทีรากจะมี H เป็ นส่วนใหญ่ ซึงได้มาจากคาร์บอนไดออกไซด์ในการ ่ หายใจของเซลล์ทเี่ ข้ารวมกับนํ้าเป็ นกรดคาร์บอนิค (H2CO3) แล้วแตกตัวให้ H+ ยึดเกาะ กับผิวรากและเคลื่อนทีในวงจํากัด บริเวณผิวรากจึงมีแรงดึงดูดระหว่างประจุบวกกับประจุ ่ ่ ลบ และสามารถแลกเปลียนไอออนกันได้ เช่น ในสารละลายทีม ี K+ มาก แล้วเคลื่อนทีเข้า ่ ่ + + ี ่ ่ี ิ ไปอยู่บริเวณราก K ก็มโอกาสทีจะถูกจับไว้ทผวรากได้เช่นเดียวกับ H และโอกาสที่ H+ + + ่ และ K จะหลุดออกจากรากก็มเี ท่ากัน ถ้า H หลุดออก K+ ก็สามารถจะเข้าไปแทนทีได้ เช่นกัน ส่วนการแลกเปลียนโดยการดูดประจุลบ เกิดจากการเข้าแทนที่ OH- หรือ HCO3่ ซึงมีอยูในรากเป็ นส่วนใหญ่ การแลกเปลียนไอออนทําให้ pH ของนํ้าในดินเปลียนแปลงไป ่ ่ ่ ่ ด้วย ถ้ามีการแลกเปลียนประจุบวกมาก pH ของนํ้าในดินจะเป็ นกรดมากขึน ่ ้ 3. กระบวนการแอกทีฟ แอพซอพชัน และการสะสมไอออน (Active Absorption and Ion Accumulation) การดูดเกลือแร่โดยวิธี Active Absorption ส่วนใหญ่เกิดทีบริเวณ ่ เนื้อเยื่อเจริญของราก ซึ่งเป็ น บริเวณที่มอตราการหายใจสูง มีพลังงานมาก จึงดูดและ ี ั สะสมเกลือแร่มาก การลําเลียงสารอาหารของพืช (Translocation of Nutrients) พืชทําการสังเคราะห์ดวยแสงทีใบได้น้ํ าตาล แล้วเปลียนเป็ นแป้ง ส่งไปตามโฟลเอม ้ ่ ่ ไปเลียงส่วนต่าง ๆ ของพืช ้ การทดลองทีแสดงว่าพืชลําเลียงอาหารไปตามท่อโฟลเอม อาจทําได้โดยการควัน ่ ่ และลอกเปลือกของลําต้นพืชใบเลี้ยงคู่ออก ทิ้งไว้ 2-3 สัปดาห์ จะสังเกตเห็นบริเวณรอย ่ ควันตอนบนโปงออก เนื่องจากอาหารทีลาเลียงลงมาจากยอดมาสะสมอยูมาก ่ ่ํ ่ CU 474 417
6.
ราบิโดและเบอร์ (Rabideau &
Burr) ทําการทดลองโดยให้ C13O2 กับใบพืช 4 กลุ่ ม ที่ทํ า ลายโฟลเอมของลํ า ต้น บริเ วณต่ า ง ๆ ด้ว ยไอนํ้ า ร้อ นหรือ ขี้ผ้ึง ร้อ น ต้น ที่ 1 ไม่ทําลายโฟลเอม ต้นที่ 2 ทําลายโฟลเอมบริเวณเหนือใบทดลอง ต้นที่ 3 ทําลายโฟลเอม บริเวณใต้ใบทดลอง และต้นที่ 4 ทําลายโฟลเอมบริเวณเหนือและใต้ใบทดลอง เมือให้ C13O2 กับใบพืชในระยะเวลาหนึ่ง แล้วตรวจหา C13 ในโมเลกุลของนํ้าตาล ่ ผลการทดลอง พบว่า ต้นที่ 1 พบทุกส่วนของลําต้น ต้นที่ 2 พบเฉพาะส่วนทีอยูใต้รอยโฟลเอมทีถูกทําลาย ่ ่ ่ ต้นที่ 3 พบเฉพาะลําต้นเหนือรอยโฟลเอมทีถูกทําลาย ่ ต้นที่ 4 พบเฉพาะบริเวณลําต้นทีอยู่ระหว่างรอยโฟลเอมทีถูกทําลาย แสดงว่า ่ ่ โฟลเอมเป็ นท่อสําหรับลําเลียงสารอาหารจากใบไปส่วนต่าง ๆ ของลําต้น รูปที่ 10.1.2 การทดลองการลําเลียงนํ้าตาลในท่อลําเลียงอาหารของพืช กลไกการลําเลียงสารอาหารของพืช สมมติฐ านที่อ ธิบ ายกลไกการลํา เลีย งสารอาหารของพืช ที่ย อมรับ กัน ทัวไปใน ่ ั ั ปจจุบน คือ Mass Flow Hypothesis ซึงอธิบายว่าสารอาหารถูกลําเลียงไปในท่อโฟลเอม ่ เกิ ด จากการผลั ก ดั น โดยความแตกต่ า งของความดั น ภายในท่ อ สมมติ ฐ านนี้ ม ี นักวิทยาศาสตร์ได้ทําการศึกษาวิจยเพื่อสนับสนุ น ดังเช่น M.H Zimmerman ได้สงเกต ั ั การใช้ปากของเพลียอ่อนเจาะเข้าไปในท่อโฟลเอมของพืช เพื่อดูดของเหลวจากท่อจนล้น ้ ออกมาทางก้น (Honen dew) การวิเคราะห์องค์ประกอบทางเคมีของของเหลวทีไม่ผ่าน ่ ลําตัวของเพลียน่ าจะเป็ นสิงทีน่าเชื่อถือมากกว่า เขาจึงตัดส่วนของลําตัวเพลียโดยการทํา ้ ่ ่ ้ ให้สลบด้วยแก๊สคาร์บอนไดออกไซด์และตัดตัวเพลี้ยด้วยแสงเลเซอร์ ของเหลวที่ได้จาก ่ ปากเพลียพบว่า ส่วนใหญ่เป็ นนํ้าตาลซูโครส ดังนันถ้าให้ 14CO2 ทางใบกับพืชเพือทําการ ้ ้ ้ สังเคราะห์ดวยแสง ซึงจะได้ซูโครสทีม ี 14C เป็ นองค์ประกอบ และเมื่อให้เพลียอ่อนแทง ้ ่ ่ 418 CU 474
7.
ปากในโฟลเอมตําแหน่ งต่าง ๆ
ก็จะหาอัตราการเคลื่อนที่ของนํ้ าตาลในโฟลเอมได้ จาก การทดลอง พบว่า นํ้าตาลในโฟลเอมสามารถเคลื่อนทีดวยอัตราประมาณ 100 เซนติเมตร ่ ้ ต่อ ชัวโมง จากผลการทดลองนี้ทําให้คดได้ว่าการเคลื่อนทีของนํ้าตาลในโฟลเอมคงไม่ใช่ ่ ิ ่ กระบวนการแพร่ธรรมดา ต้องมีพลังงานเข้ามาเกียวข้องอยูดวย ่ ่ ้ ในทฤษฎี Mass Flow ซึงเสนอโดย Ernst Munch ในปี ค.ศ. 1930 อธิบายการไหล ่ ของของเหลวในเซลล์ลําเลียงอาหารว่ามีแรงผลักดันทีเกิดจากความแตกต่างของความดัน ่ ระหว่างแหล่งสร้าง (Source) กับแหล่งเก็บ (Sink) ซึงความแตกต่างของความดันเกิดจาก ่ การลําเลียงอาหารเข้าสู่โฟลเอมที่แหล่งสร้าง และการลําเลียงอาหารออกจากโฟลเอมที่ แหล่งเก็บ พืชจะใช้พลังงานในการลําเลียงอาหารเข้าสู่โฟลเอมทีแหล่งสร้าง ทําให้ความ ่ เข้มข้นของสารละลายในโฟลเอมเพิมมากขึน ความเข้มข้นของนํ้าในเซลล์โฟลเอมจึงลดลง ่ ้ นํ้ า จากเซลล์ไ ซเลมข้า งเคียงจึงแพร่เ ข้า สู่เซลล์โ ฟลเอม ทํา ให้เ ซลล์ม ีแรงดัน เต่ง (T.P) เพิมขึ้น ในขณะเดียวกันการลําเลียงอาหารออกจากโฟลเอมไปที่เซลล์แหล่งเก็บ (Sink ่ Cell) ทําให้ความเข้มข้นของนํ้าในเซลล์โฟลเอมเพิมมากขึน เนื่องจากสารละลายมีความ ่ ้ เข้มข้นลดลง ทําให้น้ํ าแพร่ไปยังเซลล์ไซเลมทีอยู่ขางเคียง แรงดันเต่งในโฟลเอมจึงลดลง ่ ้ แรงดันเต่งทีแหล่งสร้างกับทีแหล่งเก็บจึงแตกต่างกันทําให้เกิดการแพร่ของสารจากเซลล์ ่ ่ แหล่งสร้างมายังเซลล์แหล่งเก็บได้ ซึงไม่ตองใช้พลังงาน ่ ้ รูปที่ 10.1.3 การลําเลียงสารอาหารจากแหล่งสร้างมายังแหล่งเก็บในโฟลเอม (จาก Campbell et al, 2003 และ Taiz and Zeiger, 2002) CU 474 419
8.
แม้ว่าการลําเลียงอาหารจะใช้กลไกความแตกต่างของความดันเป็ นแรงผลักดันให้ สารละลายในโฟลเอม เคลื่อนทีเป็
นระยะทางไกลแบบไม่ใช้พลังงาน (Passive) แต่พชก็ยง ่ ื ั ต้องใช้พลังงาน (Active) ในการนําสารละลายเข้าและออกจากโฟลเอมในปริมาณมากเพื่อ สร้างความแตกต่างของความดันที่จะเป็ นแรงผลักดันในการลําเลียงจากใบที่เป็ นแหล่ง สร้างไปยังรากซึงเป็ นแหล่งเก็บ ่ แต่มขอสงสัยว่าในโฟลเอมทีเป็ น Sieve Tube เป็ นเซลล์ทมชวตบริเวณแผ่นตะแกรง ี ้ ่ ่ี ี ี ิ (Sieve Plate) ยังมีสายไซโตพลาสซึมทีจะขัดขวางการลําเลียงสาร จึงไม่น่าจะผ่านได้เร็ว ่ นอกจากนี้ยงพบว่าบางครัง Turgor Pressure ในเซลล์รากก็ไม่ได้มค่าน้อยไปกว่าใบ จึงไม่ ั ้ ี ั เป็ นปจจัยทีจะทําให้เกิดแรงผลักดันทีจะลําเลียงสารได้รวดเร็ว และยังอาจเป็ นการลําเลียง ่ ่ จากรากไปที่ใ บก็ไ ด้ เพราะพบว่า การลํ า เลีย งสารอาหารของพืช มีท ง สองทิศ ทาง คือ ั้ ลําเลียงขึนและลง ้ นักวิทยาศาสตร์ได้ศึกษาวิจย เพื่ออธิบายข้อสงสัย เหล่า นี้ เพื่อสนับสนุ นทฤษฎี ั Mass Flow ดังนี้ 1. Sieve Tube Member มีรตะแกรง (Sieve Plate Pore) เป็ นช่องเปิ ด ไม่ได้อุด ู ตันในขณะทีเซลล์ยงมีชวต ซึงนักวิทยาศาสตร์ได้ทําการศึกษาโดยใช้กล้องจุลทรรศน์ชนิด ่ ั ีิ ่ พิเศษ (Confocal Laser Scanning Microscope) กับเทคนิคการให้สารสีเรืองแสงกับ สารละลายในโฟลเอมเพื่อติดตามการเคลื่อนที่ของสารอาหาร ทําให้ทราบว่าสารอาหาร สามารถลําเลียงผ่านรูตะแกรงของเซลล์ Sieve Tube Member ได้ 2. ไม่พบการลําเลียงสองทิศทางทีเกิดในเซลล์ท่ออาหารเดียวกัน นักวิทยาศาสตร์ ่ ทําการทดลองโดยใช้สารเคมี 2 ชนิดทําเครื่องหมายกับ 2 ใบทีตดกัน แต่ละใบมีสารเคมีท่ี ่ ิ ต่างกัน แล้วตรวจหาสารเคมีท่ีโฟลเอมของใบทังสอง อีกการทดลองหนึ่ง ผู้ทดลองทํา ้ เครืองหมายด้วยสารเคมีทขอ แล้วตรวจหาสารทีดานบนและล่าง จากผลการทดลองพบว่า ่ ่ี ้ ่ ้ พืชมีการลําเลียงสองทิศทางในโฟลเอมทีอยูกนคนละมัดของท่อลําเลียง หรือในมัดเดียวกัน ่ ่ ั แต่ต่างเซลล์กน ั 3. อัต ราการลํ า เลีย งสารอาหารในโฟลเอมไม่ ต อบสนองต่ อ การใช้พ ลัง งาน นัก วิทยาศาสตร์ทํา การทดลองโดยทํา ให้บ างส่ว นของก้า นใบได้ร บ อุ ณ หภูมท่ีต่ํา ลงถึง ั ิ ่ ี ้ 1 C อย่างรวดเร็ว เพือทําให้เอนไซม์ไม่ทํางานจึงไม่มการสร้างพลังงาน พบว่า การยับยัง การลําเลียงสารอาหารออกจากใบเป็ นเพียงระยะสัน อัตราการลําเลียงจะค่อย ๆ กลับคืนสู่ ้ 420 CU 474
9.
ระดับปกติ แม้ว่าความเย็นนี้จะยังทําให้อตราการหายใจทีกานใบยังคงลดลงน้อยกว่าปกติ ั ่ ้ 90
% ก็ตาม แสดงว่าการลําเลียงสารอาหารในโฟลเอมมีการใช้พลังงานน้อยมาก 4. ความแตกต่างของความดันมากพอทีจะผลักดันสารละลายให้เคลื่อนทีได้ใน ่ ่ ระยะใกล้ นักวิทยาศาสตร์ทดลองความดันโดยใช้เครื่องมือ Micromano Meter หรือ Pressure Transducer ต่อเข้ากับปากเพลี้ยอ่อนทีเจาะเข้าไปในโฟลเอม ซึ่งจะมีของ ่ เหลวไหลออกมา ค่าทีวดได้ถอว่าใกล้เคียงความเป็ นจริง เนื่องจากปากของเพลี้ยจะเจาะ ่ ั ื เซลล์เพีย งเซลล์เดีย วและรอบปากเพลี้ย จะไม่ม ีร อยรัว จากการทดลองวัด ความดัน ที่ ่ บริเวณแหล่งสร้าง (Source) กับแหล่งเก็บ (Sink) พบว่า แรงดันในเซลล์ใกล้เคียงแหล่ง สร้างมีมากกว่าเซลล์ใกล้เคียงแหล่งเก็บ ซึ่งจากการคํา นวณแรงดันของบริเวณทังสอง ้ นักวิทยาศาสตร์พบว่า มีความแตกต่างของแรงดันมาพอทีจะผลักดันให้มการไหลของ ่ ี ชีวมวลไปทางโฟลเอมได้ การคายนํ้าของพืช (Transpiration) นํ้าทีพชดูดเข้าไปทางราก จะถูกลําเลียงผ่านลําต้นไปทีใบเพื่อใช้ในกระบวนการเม ่ ื ่ แทบอลิซมเพียงเล็กน้อยเท่านัน นํ้าส่วนใหญ่จะถูกกําจัดออกไปในรูปของไอนํ้าทางปากใบ ึ ้ เรียกว่า การคายนํ้ า ซึ่งจะเกิดในเวลากลางวันที่พชมีการสังเคราะห์ด้วยแสง ส่วนในเวลา ื กลางคืนเป็ นเวลาทีปากใบปิ ด พืชจะกําจัดนํ้ าออกทางท่อไฮดราโทด (Hydrathode) ที่ ่ บริเวณปลายใบในรูปของหยดนํ้า เรียกว่า กัตเตชัน (Guttation) การคายนํ้าของพืชส่วนใหญ่เป็ นการแพร่ของไอนํ้า ซึงเกิดได้หลายบริเวณ ได้แก่ ่ 1. การคายนํ้าทางปากใบ (Stomatal Transpiration) เกิดมากทีสด ุ่ 2. การคายนํ้าทางผิวใบ (Cuticular Transpiration) เป็ นการคายนํ้าทีผวใบ ซึงมี ่ ิ ่ คิวติเคิล ฉาบอยู่ทผวเซลล์เอพิเดอร์มส เป็ นการคายนํ้ าได้น้อย เนื่องจากมีสารพวกไขมัน ่ี ิ ิ (Wax) เคลือบอยู่ ป้องกันการแพร่ของไอนํ้า 3. การคายนํ้ าทางรอยแตกของลําต้นทีเรียกว่า ตา (Lenticular Transpiration) ่ เกิดขึนน้อยมาก เนื่องจาก Lenticel มีน้อย และเป็ นส่วนของ Cork Cell ด้วย ้ CU 474 421
10.
กลไกการคายนํ้าของพืช ั เซลล์สปนจี (Spongy Cell)
ในชัน Mesophyl ของใบมีการจัดเรียงตัวอย่างหลวม ๆ ้ ทําให้เกิดช่องว่างระหว่างเซลล์มากและมีชองอากาศใหญ่ (Air Space) ทีอยู่ถดจากเซลล์ ่ ่ ั เอพิเดอร์มสด้านล่าง ซึงตรงกับปากใบ เซลล์ส ิ ่ ั ป น จีจ ะสัม ผัส กับ อากาศภายนอก ไอนํ้ า ใน ช่ อ งว่ า งจึง แพร่ อ อกมาข้า งนอกได้ ในเวลา กลางวันปากใบจะเปิ ด เนื่องจากเซลล์คุมมีการ สังเคราะห์ด้วยแสงได้น้ํ าตาล ทําให้เซลล์คุมมี สารละลายเข้ม ข้น กว่ า เซลล์ ข้า งเคีย ง จึง มี แรงดันออสโมซิส (O.P) เพิมขึน นํ้ าในเซลล์ ่ ้ ข้างเคียงจึงแพร่เข้าสูเซลล์คุมได้ ทําให้มแรงดัน ่ ี เต่ง (T.P.) ในเซลล์สงขึน ทําให้เซลล์เต่ง แต่ ู ้ รูปที่ 10.1.4 การปิดเปิดของปากใบ (จาก George B. Johnson, 1997) เนื่องจากผนังด้านนอกของเซลล์บางกว่าด้าน ใน จึงดันให้ผนังด้านนอกโค้งออกมากกว่าด้าน ใน ปากใบจึงเปิ ดกว้าง ไอนํ้ าในช่องอากาศจึงแพร่ออกมาด้านนอก ใบมีการคายนํ้ ามากก็ ส่งผลให้รากดูดนํ้ามากเพือชดเชยกับปริมาณนํ้าทีเสียไป ใบจึงเต่งอยูเสมอ ่ ่ ่ ปัจจัยที่ควบคุมการคายนํ้าของพืช การคายนํ้ าของพืชจะเกิดมากหรือน้อย เร็วหรือช้า ขึนอยู่กบสภาพแวดล้อมของ ้ ั พืช ได้แก่ 1. แสงสว่าง ในสภาพที่มความเข้มของแสงมาก จะมีผลทําให้ปากใบเปิ ดกว้าง ี เนื่องจากเซลล์คุมมีการสังเคราะห์ดวยแสงมากใบจึงคายนํ้ ามาก นอกจากนี้แสงสว่างยังมี ้ ผลทํ า ให้อุ ณ หภู ม ิข องบรรยากาศรอบใบสูง ทํ า ให้น้ํ า รอบ ๆ ใบกลายเป็ น ไอมากขึ้น ใบจึงคายนํ้ามาก 2. อุณหภูม ิ เมื่ออุณหภูมของบรรยากาศรอบ ๆ ใบสูง จะทําให้อุณหภูมของนํ้ าใน ิ ิ ใบสูงขึนด้วย นํ้ าจึงเปลียนสภาพเป็ นไอได้ง่ายและเร็ว ขณะเดียวกันอุณหภูมของอากาศ ้ ่ ิ ภายนอกใบสูง จะช่วยให้อากาศสะสมไอนํ้าได้มาก ไอนํ้าภายในใบจึงระเหยออกมาได้มาก 3. ความชืน ในบรรยากาศทีมความชืนตํ่า ซึงหมายถึงอากาศแห้งมีไอนํ้าน้อย เช่น ้ ่ ี ้ ่ เวลากลางวัน หรือ ฤดู ร้ อ น ความแตกต่ า งของความชื้น ในใบกับ นอกใบจะมีม าก 422 CU 474
11.
ทําให้การคายนํ้ าเกิดได้เร็วและมาก ในทางกลับกันถ้าอากาศมีความชืนสูง
เช่น ขณะฝน ้ ตกใหม่ ๆ เช้ามืด หรือฤดูฝน ใบจะคายนํ้าได้น้อยลง 4. ลม ขณะทีมลมพัดแรง จะพัดพาไอนํ้าไปจากบริเวณรอบใบ ทําให้เกิดความแห้ง ่ ี ทีรอบใบ ไอนํ้าจากในใบจึงระเหยออกมาสะสมมาก แต่ถาลมพัดแรงมากจนเป็ นพายุ ปาก ่ ้ ใบจะปิ ด การคายนํ้าจะลดลง ถ้าลมสงบไอนํ้าภายในกับภายนอกใบไม่แตกต่างกันมาก ใบ จะคายนํ้าได้ตลอดเวลา ในอัตราทีต่ากว่ามีลมพัด ่ ํ 5. ปริม าณนํ้ า ในดิน ถ้า ในดิน มีน้ํ า มาก รากจะดูด นํ้ า มาก และลํ า เลีย งไปที่ใ บ ตลอดเวลา ทําให้ใบต้องคายนํ้ามาก ในทางตรงข้ามกับสภาพดินทีแห้งแล้ง พืชดูดนํ้าน้อย ่ ใบก็จะคายนํ้าได้น้อย 6.โครงสร้างของใบ พืชแต่ละชนิดมีลกษณะโครงสร้างของใบแตกต่างกัน ซึงมักจะ ั ่ เป็ นการปรับตัวตามสภาพของแหล่งทีอยู่ดวย เช่น พืชทะเลทราย (Xerophyte) มีการ ่ ้ ปรับตัวโดยการเปลียนแปลงโครงสร้างของใบ เพื่อลดการคายนํ้า เช่น มีปากใบบุ๋มลึกเข้า ่ ไปในเนื้อใบ เพือทําให้เกิดแอ่งเล็ก ๆ ขังนํ้า เรียกว่า Shunken Stoma นํ้าในแอ่งจะทําให้ ่ บรรยากาศรอบใบอิมตัวด้วยความชืนและไอนํ้ า ใบจึงคายนํ้ าได้ยาก เป็ นการลดการคาย ่ ้ นํ้ า นอกจากนี้ยงมีการลดขนาดของใบ หรือดัดแปลงใบไปเป็ นหนาม เพื่อลดปากใบให้ ั น้อยลงหรือไม่ม ี ส่วนพืชที่อยู่ในที่ชุ่มชื้น (Hydrophyte) มักมีปากใบเรียบหรือนู นขึน ้ (Raised Stoma) และมีแผ่นใบขนาดใหญ่ ทําให้คายนํ้าได้มาก พืชบกทีขนในทีมความชืน ่ ้ึ ่ ี ้ ปานกลาง (Mesophyte) มักมีแผ่นใบปานกลาง มีการคายนํ้าแตกต่างกันขึนอยู่กบจํานวน ้ ั ปากใบของพืช การวัดการคายนํ้าของพืช พืชแต่ละชนิดมีการคายนํ้าทีแตกต่างกัน อัตราการคายนํ้าจึงไม่เท่ากัน การคายนํ้า ่ ของพืชคิดเป็ นนํ้ าหนักหรือปริมาตรต่อหน่ วยเวลา วิธวดทําได้หลายวิธี แต่วธทใช้กนมาก ีั ิ ี ่ี ั คือ วัดด้วยโปโตมิเตอร์ (Potometer Method) ซึงเป็ นเครืองมือทีประกอบด้วย หลอดแก้ว ่ ่ ่ บรรจุน้ํ ามีกงไม้เสียบกับจุกยางทีปากหลอด จากหลอดแก้วมีแขนยื่นออกไป 2 ข้าง แขน ิ่ ่ ด้า นซ้า ยติด กับ กรวยที่ม ีก๊ อ กปิ ด เปิ ด สํา หรับ เติม นํ้ า เข้า หลอดแก้ว แขนด้า นขวาเป็ น หลอดแก้วทีมรขนาดเล็กมาก (Capillary Tube) มีมาตราส่วนแบ่งเป็ นปริมาตรทีมความ ่ ีู ่ ี ละเอียด ปลายหลอดงอจุมอยูในแก้วบรรจุน้ําสี ่ ่ CU 474 423
12.
รูปที่ 10.1.5 โปโตมิเตอร์สาหรับวัดอัตราการคายนํ้าของพืช
(จาก John W. Kimball, 1965) ํ การทดลองหาอัตราการคายนํ้ าของพืช ก่อนทําการทดลองนํ าถ้วยบรรจุน้ํ าออก จากปลาย Capillary Tube ก่อน เพื่อให้เกิดฟองอากาศเข้าสู่หลอด เมื่อใบพืชคายนํ้ า ฟองอากาศจะเคลื่อนทีใช้เป็ นตัวชีวดบนมาตราส่วน เพื่อบอกปริมาตรของไอนํ้าทีพชคาย ่ ้ั ่ ื ออกไปทางใบ ต่อจากนี้จงจุมปลายหลอด Capillary ลงไปในถ้วยนํ้าสี ทีหลอดแก้วใหญ่ จะ ึ ่ ่ มีต้นไม้ทงต้น หรือ เฉพาะกิ่ง เสีย บแน่ นทะลุแผ่น จุก ยาง มีห ลอดไฟฟ้ า ให้แสงสว่า งอยู่ ั้ ภายนอก เมื่อเตรียมเครื่องมือเสร็จเรียบร้อยแล้ว เริมจับเวลาที่ฟองอากาศ เคลื่อนที่ใน ่ Capillary อ่านปริมาตรของนํ้าทีถูกดูดเข้าสูลาต้นแล้วระเหยออกทางใบ วัดพืนทีของใบพืช ่ ่ํ ้ ่ ทังหมด นํ ามาคํานวณหาอัตราการคายนํ้าของพืชต่อพืนทีผวใบ 1 ตารางหน่ วยในเวลา 1 ้ ้ ่ ิ หน่วย ตัวอย่าง ทดลองการคายนํ้ าของพืชโดยใช้โปโตมิเตอร์ทหลอดแคปพิลลารีมเส้น ่ี ี ผ่านศูนย์กลางของรูภายใน 0.8 มิลลิเมตร ยาว 30 เซนติเมตร จัดเตรียมเครื่องมือดังภาพ ในเวลา 1 นาที ฟองอากาศในหลอดเคลื่อนทีไปเป็ นระยะทาง 7 เซนติเมตร อัตราการคาย ่ นํ้าของกิงไม้ททดลองเป็ นเท่าใด ่ ่ี เส้นผ่านศูนย์กลางของรูหลอดแคปพิลลารี = 0.8 มิลลิเมตร = 0.08 เซนติเมตร พืนทีหน้าตัดของรูภายในหลอด ้ ่ = πr2 = 22 x ( 0.08 ) 2 ตารางเซนติเมตร 2 7 ปริมาตรของนํ้าในหลอดแคปพิลลารี = 7 x 22 (0.04)2 ลูกบาศก์เซนติเมตร = 0.3527ลูกบาศก์เซนติเมตร ∴ อัตราการคายนํ้าของกิงไม้ทดลอง = 0.352 ลูกบาศก์เซนติเมตร / นาที ่ 424 CU 474
13.
การคายนํ้าในรูปหยดนํ้าของพืช (Guttation) การคายนํ้าของพืชทีออกมาเป็ นหยดนํ้า
ส่วนใหญ่เกิดบริเวณปลายใบและขอบใบที่ ่ เป็ นรอยหยักแหลม และมักเกิดเนื่องจากสภาพแวดล้อมภายนอกไม่เอืออํานวยให้เกิดการ ้ คายนํ้าในรูปของไอนํ้า เช่น อากาศมีความชืนสูง อิมตัวด้วยไอนํ้า ลมสงบ อุณหภูมต่ํา และ ้ ่ ิ ไม่มแสงสว่าง ทําให้ปากใบไม่เปิ ด แต่รากมีการดูดนํ้ าตลอดเวลา นํ้ า ที่ถูกดันขึ้นมาจะ ี สินสุดทีปลายของท่อเทรคีด (Tracheid) ปลายเส้นใบ ซึงจะติดต่อกับเซลล์พาเรนไคมาที่ ้ ่ ่ จัดเรียงตัวอย่างหลวม ๆ บางช่วงบางตอนมีรูทอาจต่อเนื่องไปถึงปลายใบที่มปากใบอยู่ ่ี ี ด้ว ย ดัง นั น ในเวลากลางคืน ที่ป ากใบปิ ด ก็ย ัง มีรู เ ล็ก ๆ เปิ ด อยู่ เรีย กว่ า ไฮดรา-โทด ้ (Hydrathode) นํ้ าจะดันผ่านเซลล์พาเรนไคมามายังรูทปลายใบหรือขอบใบได้กลายเป็ น ่ี หยดนํ้า 10.2 การสังเคราะห์ด้วยแสง (Photosynthesis) อาจกล่าวได้ว่าพลังของสิงมีชวต คือ แสงอาทิตย์ทงนี้เนื่องจากพลังงานทีสงมีชวต ่ ีิ ั้ ่ ิ่ ี ิ ใช้ในการดํารงชีวตทังหมด ล้วนได้มาจากดวงอาทิตย์ทจบไว้โดยพืชและสาหร่ายผ่านทาง ิ ้ ่ี ั กระบวนการสังเคราะห์ดวยแสง ทุกชีวตจึงดํารงอยู่ได้ดวยพลังงานแสงจากดวงอาทิตย์ ใน ้ ิ ้ แต่ละวันพลังงานจากแสงอาทิตย์ทสองลงมายังพืนโลก มีจานวนมหาศาล นักวิทยาศาสตร์ ่ี ่ ้ ํ ประมาณไว้วามีถง 1 ล้านเท่าของอะตอมมิกบอมบ์ ทีนําไปทิงทีเกาะฮิโรชิมา แต่พชสีเขียว ่ ึ ่ ้ ่ ื และสาหร่า ยจับ พลังงานนี้ ไ ว้เพีย ง 1 เปอร์เ ซ็น ต์เ ท่า นัน ก็เ พีย งพอที่จ ะใช้ส ร้า งอาหาร ้ ั แบ่งปนให้กบทุกชีวตบนโลกสามารถดํารงชีพอยูได้ ั ิ ่ ภาพรวมของกระบวนการสังเคราะห์ด้วยแสง กระบวนการสังเคราะห์ดวยแสงเกิดขึนทีใบของพืชเป็ นส่วนใหญ่ ทีเซลล์ใบของพืช ้ ้ ่ ่ มีออร์แกเนลล์ทเี่ รียกว่า คลอโรพลาสต์ (Chloroplast) เป็ นแหล่งสําคัญทีทาให้กระบวนการ ่ ํ ดําเนินไปได้ตลอด ซึ่งไม่มโครงสร้า งใดในเซลล์พชที่จะทดแทนได้ นักวิทยาศาสตร์ได้ ี ื ทําการศึกษาค้นคว้าจนพบความจริงว่า กระบวนการสังเคราะห์ดวยแสงจะดําเนินไปใน 3 ้ ขัน คือ 1) จับพลังงานจากแสงอาทิตย์ 2) ใช้พลังงานสร้างสารเก็บพลังงาน คือ ATP และ ้ NADPH และ 3) ใช้ ATP เป็ นพลังงานในการสร้างโมเลกุลของพืชจาก CO2 ในอากาศ โดยมีการศึกษาปฏิกรยาการสังเคราะห์ดวยแสง ดังนี้ ิิ ้ CU 474 425
14.
ในปี ค.ศ. 1932
Robin Hill ได้ทําการแยกคลอโรพลาสต์ออกจากเซลล์ผกโขม ั นํามาทดลองปฏิกรยาการแตกตัวของนํ้ าด้วยแสง โดยผ่านแสงไปในของผสมของเกลือเฟ ิิ อริกและคลอโรพลาสต์ แล้วพบว่า เกลือเฟอริกเปลียนเป็ นเกลือเฟอรัสกับ ออกซิเจน ่ 4Fe3+ + 4H2O Light Chloroplast 4Fe2+ + 4H+ + O2 + H2O และพบว่าในปฏิกรยาที่ไม่มเกลือเฟอริกจะไม่เกิดออกซิเจน ในปฏิกิรยาที่เกลือเฟอริก ิิ ี ิ เปลี่ย นเป็ น เกลือ เฟอรัสได้ เนื่ อ งจากได้ร บ อิเ ลคตรอนจากนํ้ า ที่แ ตกตัว เมื่อ มีค ลอโรั พลาสต์ แ ละแสง ส่ ว นออกซิเ จนที่เ กิด ขึ้น ในปฏิกิร ิย าแสดงว่ า มีเ กลือ เฟอริก เป็ น ตัว ออกซิไดซ์ หรือตัวรับไฮโดรเจน (Hydrogen Acceptor) สารทีทาหน้าทีเป็ นตัวรับไฮโดรเจนอาจมีหลายชนิด เช่น Ferric Cyanide และ ่ ํ ่ Methylene Blue เป็ นต้น แต่ในพืชมี Nicotinamide Adenosine Dinucleotide Phosphate ่ ่ ่ (NADP+) ซึงอยูในรูปของออกซิแดนท์ เป็ นตัวรับไฮโดรเจน เมือรับไฮโดรเจนเข้ามาจะ เปลียนรูปเป็ น รีดกแตนท์ (Reductant) คือ NADPH ่ ั จากการทดลองนี้นกวิทยาศาสตร์จงเกิดแนวคิดว่าปฏิกรยาการสังเคราะห์ดวยแสง ั ึ ิิ ้ น่ าจะมีขนตอนการปล่อยออกซิเจนและขันตอนที่เกี่ยวข้องกับคาร์บอนไดออกไซด์ ดังที่ ั้ ้ Danial Arnon และคณะ (ค.ศ. 1651) ได้ทําการทดลองเพื่อติดตามขันตอนการ ้ + เกิดปฏิกรยาโดยผ่านแสงลงไปในสารผสมของ ADP, Pi, NADP , H2O และ Chloroplast ิิ พบว่า สารเคมีทเี่ กิดขึน คือ ATP, NADPH และ O2 ดังสมการ ้ ADP + Pi + NADP+ + H2O Light ATP + NADPH + H+ + O2 Chloroplast และพบว่าถ้าผ่านแสงลงไปในของผสมทีมเี ฉพาะ ADP, Pi และ H2O ก็จะได้ ATP เพียง ่ อย่างเดียว ซึ่งแสดงว่า การสร้าง ATP และ NADPH ย่อมขึนอยู่กบปจจัยทีให้กบ ้ ั ั ่ ั คลอโรพลาสต์ในสภาพทีตองมีแสง ่ ้ ี เมือหาการทดลองโดยให้ ATP, NADPH และ CO2 แก่คลอโรพลาสต์โดยไม่มแสง ่ ผลทีเกิดขึน คือ โมเลกุลนํ้าตาล ดังสมการ ่ ้ CO2 + ATP + NADPH 426 Chloroplast Sugar + ADP + Pi + NADP+ CU 474
15.
แสดงว่าในขันตอนการสร้างโมเลกุลของพืช คือ นํ้าตาลของคลอโรพลาสต์ตองมี ้ ้ ั ํ ปจจัย
คือ CO2, ATP และ NADPH โดยไม่จาเป็ นต้องมีแสง จากการทดลองอาจสรุปได้ว่า กระบวนการสังเคราะห์ดวยแสงของพืชในขันตอนที่ ้ ้ มีการจับพลังงานจากแสงอาทิตย์นําไปใช้สร้างสารเก็บพลังงาน คือ ATP และ NADPH จะ เกิดในสภาวะทีมแสง เป็ นปฏิกรยาทีตองใช้แสง (Light Dependent Reaction) ซึงเป็ น ่ ี ิิ ่ ้ ่ ขันตอนแรก ส่วนขันตอนที่ 2 เป็ นการนํ าพลังงานจากแสงในรูปของ ATP มาใช้เปลียน ้ ้ ่ CO2 ให้เป็ นโมเลกุลนํ้ าตาลจะเกิดในสภาวะที่ไม่ต้องมีแสงมาเกี่ยวข้องหรือมืด เป็ น ปฏิกรยาทีไม่ตองใช้แสง (Light - Independent or Dark Reaction) ิิ ่ ้ โดยภาพรวมของกระบวนการสังเคราะห์ดวยแสง อาจสรุปได้จากสมการ ดังนี้ ้ 6CO2 + 12H2O + Light energy Carbon Water dioxide C6H12O6 + 6H2O + 6O2 Glucose Water Oxygen คลอโรพลาสต์ (Chloroplast) คลอโรพลาสต์ (Chloroplast) คือ ออร์แกเนลล์ของพืชทีเป็ นแหล่งเกิดกระบวนการ ่ สังเคราะห์ดวยแสงของพืช โครงสร้างประกอบด้วย เยื่อ 2 ชัน เยื่อชันนอกเรียบ เยื่อชันใน ้ ้ ้ ้ ยื่นเข้าไปข้างใน มีลกษณะเป็ นถุงกลมแบนคล้ายเหรียญ เรียกว่า ไทลาคอยด์ (Thylakoid) ั ไทลาคอยด์จานวนมากมีการจัดเรียงตัวโดยวางซ้อนกันเป็ นตัง เรียกว่า กรานุ่ม (Granum) ํ ้ แต่ละคลอโรพลาสต์ม ี 40-60 กรานุ่ ม แต่ละกรานุ่ มเชื่อมต่อกันด้วยโครงสร้างที่เรียกว่า สโตรมา ลาเมลลา (Stroma Lamella) ซึงจะพบเฉพาะในพืชชันสูง รอบ ๆ เยื่อไทลาคอยด์ ่ ้ เป็ นสารกึงเหลว (Semifluid) เรียกว่า สโตรมา (Stroma) ประกอบด้วย เอนไซม์ DNA ่ RNA และไรโบโซม ทําให้คลอโรพลาสต์สามารถสร้างโปรตีนของตัวเองได้ในบางกรณี และสามารถแบ่งตัว (Division) ได้ ไทลาคอยด์เป็ นทีอยู่ของสารสีพวกคลอโรฟิลและสารสี ่ ประกอบอื่น ๆ โดยที่ผ ิว ของไทลาคอยด์ม ีก ารจัด กลุ่ ม สารสีช นิ ด เดีย วกัน เป็ น ร่ า งแห เรียกว่า ระบบแสง (Photosystem) CU 474 427
16.
รูปที่ 10.2.1 โครงสร้างของคลอโรพลาสต์ (จาก
George B. Johnson 1995 The Living World หน้า 92, 93) สารสี (Pigment) สารสี (Pigment) คือ โมเลกุลของสารทีสามารถดูดพลังงานแสงได้ มีหลายชนิด ่ แต่ละชนิดมีความสามารถดูดพลังงานแสงในช่วงคลื่นทีแตกต่างกัน แสงทีตาคนมองเห็น ่ ่ กับแสงทีตาสัตว์มองเห็นอาจเป็ นแสงในช่วงคลื่นทีต่างกัน เนื่องจากตาของคนและสัตว์ม ี ่ ่ สารสีทแตกต่างกันนันเอง แสงทีตาคนมองเห็น (Visible Light) คือ แสงทีมความยาวคลื่น ่ี ่ ่ ่ ี ทีสารสีในตา คือ เรตินล (Retinal) สามารถดูดได้ ซึงอยู่ระหว่าง 380 นาโนเมตร (สีมวง) ่ ั ่ ่ ถึ ง 750 นาโนเมตร (สีแ ดง) ในขณะที่ส ัต ว์ อ่ื น ๆ ใช้ ส ารสีใ นการมองเห็ น แถบสี (Spectrum) ทีแตกต่างกัน เช่น ตาแมลงมีสารสีทดดแสงช่วงคลื่นสันกว่าเรตินลของคน ซึง ่ ่ี ู ้ ั ่ เป็ นสาเหตุททาให้ผงสามารถมองเห็นแสง Ultraviolet (UV) ได้ในขณะทีตาของคนมองไม่ ่ี ํ ้ึ ่ เห็น แต่แมลงก็ไม่สามารถมองเห็นแสงสีแดงเหมือนอย่างทีคนเห็น สารสีทสาคัญทีพชใช้ ่ ่ี ํ ่ ื ดูดพลังงานแสง คือ คลอโรฟิ ล เป็ นสารสีทแม้ว่าจะดูดชนิดของโฟตอนได้น้อยกว่าเรตินัล ่ี ของคน แต่กมประสิทธิภาพในการจับโฟตอนได้มากกว่า เนื่องจากโมเลกุลมีแมกเนเซียม ็ ี เป็ นองค์ประกอบ ซึงใช้จบกับโฟตอน ่ ั 428 CU 474
17.
สารสีทใช้ในกระบวนการสังเคราะห์ดวยแสงในสิงมีชวตชนิดต่าง ๆ มีหลายชนิด ่ี ้ ่
ีิ ดังตารางต่อไปนี้ ตารางที่ 10.2.1 สารสีในกระบวนการสังเคราะห์ดวยแสงของสิงมีชวต ้ ่ ีิ ชนิดของสิงมีชวต ่ ีิ ยูคาริโอต มอสส์ เฟิรน ์ พืชดอก สาหร่ายสีเขียว สาหร่ายสีน้ําตาล สาหร่ายสีแดง โปรคาริโอต สาหร่าย สีเขียวแกมนํ้าเงิน ซัลเฟอร์ เพอร์เพิล แบคทีเรีย กรีน แบคทีเรีย คลอโรฟิล a b c d a แบคเทอริโอคลอโรฟิล คาโร b c d e g ตินอยด์ ไฟโค บิลน ิ + + - - + - + + - + - + + - - + + + + + + - - + + + + + - - + หรือ + + - + หรือ + หรือ + - คลอโรฟิ ล (Chlorophyll) คลอโรฟิ ล (Chlorophyll) เป็ นสารสีท่ีมสเขียว เปลี่ยนแปลงมาจากโปรโตี ี คลอโรฟิล (Protochlorophyll) เมือถูกแสง โมเลกุลมีโลหะแมกนีเซียม (Mg) ทีจบอยู่กบ ่ ่ั ั ไนโตรเจน อยู่กลางโมเลกุ ลที่เป็ นวงแหวนคาร์บอน และมีส่วนหางที่เป็ นสารประกอบ คาร์บอนกับไฮโดรเจน (Hydrocarbon Tail) คลอโรฟิ ลมี 4 ชนิด คือ คลอโรฟิ ล เอ บี ซี และดี แต่ละชนิดมีโครงสร้างเคมีต่างกันทีกลุ่มด้านข้างเท่านัน และสามารถดูดแสงในช่วง ่ ้ ั คลื่นทีต่างกัน โมเลกุลของคลอโรฟิ ลจะฝงตัวอยู่บนโมเลกุลโปรตีนทีเป็ นองค์ประกอบของ ่ ่ เยือไทลาคอยด์ ่ CU 474 429
18.
รูปที่ 10.2.2 โครงสร้างโมลกุลของคลอโรฟิล
เอ และ บี (ก) และแบบจําลองตําแหน่งของคลอโรฟิลบนโปรตีน ทีเยือไทลาคอยด์ (ข) (จาก Purves et al. , 2003. Life : The Science of Biology, 7th Edition ในโครงการ ่ ่ ตําราวิทยาศาสตร์และคณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 2 หน้า 64) แคโรทีนอยด์ (Carotenoid) แคโรทีนอยด์ เป็ นสารสีทมสสมแดง ไม่ละลายนํ้า มี 2 ชนิด คือ แคโรทีน (Carotene) ่ี ี ี ้ มีสสม และแซนโทฟิล (Xanthophyll) มีสเี หลือง ซึงมักอยู่รวมกันกับคลอโรฟิลในคลอโรพ ี้ ่ ่ ลาสต์ แต่พชบางชนิดมีแคโรที นอยด์อยู่ในโครโมพลาสต์ (Chromoplast) เช่น แครอท ื มะเขือเทศ และผลไม้ทมสเี หลือง เป็ นต้น แคโรทีนอยด์เป็ นสารสีประกอบ มีหน้าทีช่วยดูด ่ี ี ่ พลังงานแสงช่วงคลื่น 400-500 นาโนเมตร ส่งให้คลอโรฟิ ลเอ ซึ่งเป็ นช่วงคลื่นที่แคโรที นอยด์ดูดได้ดทสุดในขณะทีคลอโรฟิ ล เอ ดูดได้น้อย นอกจากนี้ยงช่วยป้องกันไม่ให้คลอ ี ่ี ่ ั โรฟิลถูกทําลายจากแสงในปฏิกรยาโฟโตออกซิเดชัน (Photo Oxidation) อีกด้วย ิิ ไฟโคบิ ลิน (Phycobilin) ไฟโคบิลน เป็ นสารสีทพบเฉพาะในสาหร่ายสีแดงและสาหร่ายสีเขียวแกมนํ้าเงิน มี ิ ่ี 2 ชนิด คือ ไฟโคอีรทน (Phycoerythin) มีสารสีแดง ดูดแสงช่วงคลื่น 495-565 นาโนเมตร ซึง ี ิ ่ เป็ นสีเขียว พบในสาหร่ายสีแดง อีกชนิดหนึ่ง คือ ไฟโคไซยานิน (Phycocyanin) เป็ นสาร 430 CU 474
19.
สีน้ําเงิน ดูดแสงช่วงคลืน 550-615
นาโนเมตร พบในสาหร่ายสีเขียวแกมนํ้าเงิน ไฟโคบิลน ่ ิ เป็ นสารสีประกอบโดยทําหน้าทีดดพลังงานแสงส่งให้กบคลอโรฟิล เอ ่ ู ั แบคทีริโอคลอโรฟิ ล (Bacteriochlorophyll) แบคทีรโอคลอโรฟิ ล เป็ นสารสีทพบในแบคทีเรียทีสงเคราะห์ดวยแสงได้เป็ นสารสี ิ ่ี ่ ั ้ ทีดูดพลังงานแสงได้ดในช่วงคลื่น 358 ซึงเป็ น Ultraviolet และ 772 นาโนเมตร ซึงเป็ น ่ ี ่ ่ Infrared นอกจากนี้แบคทีเรียพวก Green-Sulfur Bacteria เช่น คลอโรเบียม (Chlorobium) สามารถดูดพลังงานแสงได้ดทชวงคลืน 650 นาโนเมตร และ 660 นาโนเมตร ี ่ี ่ ่ รูปที่ 10.2.3 การดูดแสงของโมเลกุลสารสีชนิดต่าง ๆ ทีความยาวคลื่นต่าง ๆ ่ th (จาก Purves et al. , 2003. Life : The Science of Biology, 7 Edition ในโครงการตําราวิทยาศาสตร์และ คณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 2 หน้า 65) การจับพลังงานจากแสงอาทิ ตย์ แสงมีคุณสมบัตเิ ป็ นทังคลืนและอนุภาค พลังงานแสงอยู่ในรูปของอนุ ภาคทีเรียกว่า ้ ่ ่ โฟตอน (Photon) แต่ละโฟตอนมีพลังงานทีเรียกว่า ควอนตัม (Quantum) ซึงแปรผกผัน ่ ่ กับความยาวคลื่นแสง คือ เมื่อแสงมีความยาวคลื่นสันจะมีระดับพลังงานสูง พลังงานแสง ้ คํานวณได้จากสมการ E = hv (Quantum) เมือ h เป็ นค่าคงที่ (Planck’s Constant ; h = ่ -34 6.626 x 10 Js) แสงอาทิต ย์ บ างช่ ว งคลื่น มีโ ฟตอนมาก ทํ า ให้ ม ีร ะดับ พลัง งานสู ง มาก เช่ น รังสีเอกซ (x-rays) และแสงอัลตราไวโอเลต (UV) บางช่วงคลื่นทีมโฟตอนน้อยจะมีระดับ ่ ี CU 474 431
20.
พลังงานตํ่า เช่น คลื่นวิทยุมโฟตอนน้อยมาก
โดยทัวไปแสงทีมพลังงานสูงจะมีความยาว ี ่ ่ ี คลื่นแสงสันกว่าแสงทีมพลังงานตํ่า ตาของคนสามารถรับแสงทีมโฟตอนอยู่ในระดับปาน ้ ่ ี ่ ี กลาง เรียกว่า แสงทีมองเห็น (Visible Light) เนื่องจากตาของคนสามารถดูดโฟตอนของ ่ แสงชนิดนัน ๆ ได้ ในพืชแม้จะดูดโฟตอนของแสงได้มากชนิดกว่าตาของคน แต่ชวงคลื่นที่ ้ ่ พืชดูดเป็ นหลัก คือ สีน้ํ าเงินและสีแดง และสะท้อนกลับสีเขียว การทีตาของเราเห็นใบไม้ ่ เป็ นสีเขียว เป็ นเพราะใบของพืชจะดูดคลื่นแสงทังหมด ยกเว้นคลื่นแสง 500-600 นาโน ้ เมตร ซึ่งเป็ นสีเขียวจะสะท้อนกลับ และสารสีท่ตาจะดูดเอาไว้ ทําให้สมองรับรู้ว่าเป็ นสี ี เขียว รูปที่ 10.2.4 สารสีของตาดูดแสงสีเขียวทีสะท้อนกลับจากใบพืช ่ (จาก George B. Johnson, 1995. The Living World หน้า 94) แสงทีตามองเห็นเป็ นสัดส่วนทีน้อยมากของโฟตอนในแสงอาทิตย์ เรียกช่วง โฟตอน ่ ่ ของแสงทีเห็นว่า Electromagnetic Spectrum แสงทีมความยาวคลื่นสันทีสด และโฟตอน ่ ่ ี ้ ุ่ มีพลังงานสูงทีสด คือ รังสีแกมมา (Gamma-rays) มีชวงคลื่นน้อยกว่า 1 นาโนเมตร และ ุ่ ่ คลืนแสงทีมพลังงานตํ่าทีสดมีความยาวคลืนเป็ นพันเมตรได้แก่ คลืนวิทยุ ่ ่ ี ุ่ ่ ่ 432 CU 474
21.
รูปที่ 10.2.5 โฟตอนของพลังงานทีแตกต่างกัน
: Electromagnetic Spectrum ่ (จาก George B. Johnson, 1995. The Living World หน้า 94) การจับพลังงานแสงของสารสีเกิดจากการเปลี่ยนแปลงระดับพลังงานของอิเลคตรอน ในโมเลกุลของสารสี เมือได้รบพลังงานแสง อิเลคตรอนทีเคลื่อนทีในวงโคจรรอบนิวเคลียส ่ ั ่ ่ ของอะตอม เป็ นสิงบ่งบอกถึงระดับพลังงานในโมเลกุลของสาร กล่าวคือ ในสภาวะปกติ ่ (Ground State) อิเลคตรอนจะรักษาระยะห่างระหว่างนิวเคลียสกับอิเลคตรอนให้คงที่ ซึง ่ จะมีค่าพลังงานน้อย เมื่อโมเลกุลของสารสีได้รบแสงสว่างโฟตอนจะพุ่งชนอิเลคตรอนพร้อม ั กับถ่ายทอดพลังงานให้ ทําให้อเลคตรอนเคลื่อนทีเร็วขึนและห่างออกจากวงโคจร โมเลกุล ิ ่ ้ ของสารสีอยู่ในภาวะตื่นตัว (Excited State) กลายเป็ นสารสีทถูกกระตุ้น (Activate ่ี Pigment) และทํางานได้โดยมีพลังงานเพิมขึน ่ ้ Pigment Molecule (P) + Quantum (hv) Activate Pigment (P+) สารสีท่ีอ ยู่ใ นสภาวะตื่น ตัว นี้ ไ ม่อ ยู่ต ว จะกลับ เข้า สู่ส ภาวะปกติโ ดยอิเ ลคตรอน ั ปลดปล่อยพลังงานทีดดไว้ออก เพือเคลือนทีกลับเข้าสูวงโคจรเดิม ซึงมีระดับพลังงานปกติ ่ ู ่ ่ ่ ่ ่ สารสีสามารถปลดปล่อยพลังงานทีได้รบมาจากแสงโดยการเรืองแสง (Fluorescence) เช่น ่ ั คลอโรฟิ ล เอ ดูดแสงทีความยาวคลื่น 660 มิลลิไมโครเมตร จะเปล่งแสงในช่วงความยาว ่ คลื่นมากกว่า 700 มิลลิไมโครเมตร ซึ่งเป็ นช่วงคลื่นสีแดง ทังนี้เนื่องจากแสงที่ดูด ไว้ม ี ้ พลังงานสูงกว่าแสงที่เปล่งออกมา พลังงานบางส่วนสูญเสียไปในรูปของความร้อน แสง และกระบวนการทางเคมี นอกจากนี้อิเลคตรอนที่อยู่ในภาวะตื่นตัว อาจหลุดออกจาก CU 474 433
22.
โมเลกุลของสารสี ก่อนจะเข้าสูวงโคจรเดิม โดยมีสารประกอบหลายชนิดเป็
นตัวรับอิเลคตรอนที่ ่ จะถ่ายทอดกันไปเป็ นทอด ๆ เรียกว่า ตัวรับอิเลคตรอน (Electron Acceptor) ซึงเป็ นสาร ่ พวก Coenzyme ได้แก่ Ferridoxin NADP และ Cytochrome สารสีส่วนใหญ่ทําหน้าที่รบพลังงานแสงและส่งต่อให้โมเลกุลอื่นถ่ายทอดกันเป็ น ั ทอด ๆ ไปจนถึงโมเลกุลทีเป็ นศูนย์กลางการรับแสง (Reaction Center) ดังนัน สารสีสวน ่ ้ ่ ใหญ่จงเปรียบเสมือนกับตัวรับสัญญาณ (Antenna) ของวิทยุโทรทัศน์ทสงภาพและเสียงไป ึ ่ี ่ ทีเครืองรับ ่ ่ รูปที่ 10.2.6 การรับและถ่ายทอดพลังงานของสารสีไปยังศูนย์กลางปฏิกรยาเคมี ิิ th (จาก Purves et al. , 2003. Life : The Science of Biology, 7 Edition ในโครงการตําราวิทยาศาสตร์และ คณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 2 หน้า 67) สารสีในคลอโรพลาสต์ จะรวมกันเป็ นหน่วยสังเคราะห์แสง (Photosynthetic Unit หรือ Quantasome) ย่อย ๆ บนไทลาคอยด์ของกรานุ่มแต่ละหน่วยประกอบด้วยคลอโรฟิล 400-600 โมเลกุล มีศูนย์กลางการรับแสง (Reaction Center) 2 ระบบ คือ ระบบแสง I (Photosystem I, PS I) และระบบแสง II (Photosystem II, PS II) PS I มีสารสีระบบ I (Pigment System I) เป็ นตัวรับแสง ประกอบด้วย คลอโรฟิล เอ ชนิดที่ดูดพลังงานแสงความยาวคลื่น 683 นาโนเมตร กับ คลอโรฟิ ล เอพิเศษ ที่ดูด พลังงานแสงความยาวคลื่น 700 นาโนเมตร เรียกสารสีระบบ I ว่า P 700 ซึงหมายถึง ่ โมเลกุลคลอโรฟิลทีทาหน้าทีเป็ นศูนย์กลางการรับแสงของระบบแสง I ่ ํ ่ PS II มีสารสีระบบ II (Pigment System II) เป็ นตัวรับแสง ประกอบด้วย คลอโรฟิล เอ ชนิดทีดูดพลังงานแสงความยาวคลื่น 673 นาโนเมตร กับคลอโรฟิ ล บี เป็ นส่วนใหญ่ และ ่ สารสีอ่น ๆ เช่น แคโรทีนอยด์ และไฟโคบิลน เรียกสารสีระบบ II ว่า P 680 ซึงหมายถึง ื ิ ่ 434 CU 474
23.
โมเลกุลคลอโรฟิลทีทาหน้าทีเป็ นศูนย์กลางการรับแสง ของระบบแสง
II (P เป็ นอักษรย่อ ่ ํ ่ ของ Pigment และใช้ค่าความยาวคลื่นแสงทีดดพลังงานแสงได้มากทีสด เป็ นค่าของระบบ ่ ู ุ่ สารสี) PS II ดูดพลังงานแสงคลื่นสันกว่า PS I ในแต่ละระบบแสงมีคลอโรฟิล ประมาณ ้ 200-300 โมเลกุล ปฏิกิริยาเคมีในกระบวนการสังเคราะห์ด้วยแสง กระบวนการสังเคราะห์ดวยแสง มีปฏิกรยาเคมีแบ่งเป็ น 2 ตอนทีเกิดขึนอย่างต่อเนื่อง ้ ิิ ่ ้ คือ ปฏิกรยาทีตองใช้แสง และปฏิกรยาทีไม่ตองใช้แสง ดังนี้ ิิ ่ ้ ิิ ่ ้ ปฏิ กิริยาที่ต้องใช้แสง (Light Dependent Reaction) ปฏิกรยาทีตองใช้แสง เป็ นกระบวนการทีพชตรึง (Fixation) พลังงาน เพือเปลียนเป็ น ิิ ่ ้ ่ ื ่ ่ พลังงานเคมีเก็บอยู่ในสารประกอบ ATP และ NADPH สําหรับใช้ในกระบวนการตรึง CO2 ในปฏิกรยาทีไม่ตองใช้แสง ATP เป็ นสารทีมพนธะเคมีพลังงานสูง (High Energy Bond) ิิ ่ ้ ่ ี ั ทีเซลล์ใช้ในกิจกรรมสําคัญ 4 อย่าง ได้แก่ ในปฏิกรยาเคมีตามทีเซลล์ตองการ การขนส่ง ่ ิิ ่ ้ สารข้ามเยื่อเซลล์หรือออร์แกเนลล์ และการเคลื่อนที่ขององค์ประกอบภายในเซลล์ ส่วน ิิ ่ ้ NADPH เป็ นสารทีพชใช้ในการตรึงและรีดวส์ CO2 ในปฏิกรยาทีไม่ตองใช้แสง ่ ื ิ ปฏิกรยาทีตองใช้แสง ประกอบด้วย 2 ปฏิกรยา คือ Photolysis หรือ Hill Reaction ิิ ่ ้ ิิ เป็ นปฏิกรยาทีพลังงานแสงทําให้น้ําแตกตัว และมีการเคลื่อนย้ายอิเลคตรอนจากโมเลกุล ิิ ่ ่ ิิ ของนํ้าไปรีดวส์ NADP+ เพือสร้าง NADPH อีกปฏิกรยา คือ Photophosphorylation เป็ น ิ ปฏิกรยาทีพชนําพลังงานแสงไปใช้สร้าง ATP โดยการเพิมหมูฟอสเฟต (Pi) ให้กบ ADP ิิ ่ ื ่ ่ ั กระบวนการตรึงพลังงานแสงบนเยื่อไทลาคอยด์ต้อ งอาศัยสารที่ป ระกอบด้ว ย โปรตีน 4 กลุ่ม ได้แก่ ระบบแสง I ระบบแสง II ATP Synthase ซึงเป็ นเอนไซม์ ในปฏิกรยาการ ่ ิิ สร้าง ATP และสารทีทาหน้าทีเป็ นตัวรับและถ่ายทอดอิเลคตรอน ได้แก่ Cytochrome b6 ่ ํ ่ Cytochrom f และ Plastocyanin การถ่ายทอดอิเลคตรอนในปฏิกรยาทีตองใช้แสง เกิดได้ 2 แบบ ได้แก่ ิิ ่ ้ 1. การถ่ายทอดอิเลคตรอนเป็ นวัฎจักร (Cyclic Electron Transfer) เป็ นการ ถ่ายทอดอิเลคตรอนในระบบแสง I เพียงระบบเดียว เมือ P 700 ถูกกระตุนด้วยพลังงาน ่ ้ แสง 2 โฟตอน (hv) 2 อิเลคตรอนของสารสีจะอยู่ในภาวะตื่นตัว ทําให้ค่าศักย์ไฟฟ้าเปลียน ่ CU 474 435
24.
จาก +0.4 ev
ไปเป็ น -0.6 ev ซึงเป็ นค่าทีใกล้เคียงกับสารรับอิเลคตรอนชนิดแรกทีไม่ ่ ่ ่ ทราบแน่ ชดว่าเป็ นสารอะไร (x) คลอโรฟิ ลจึงถ่ายทอดอิเลคตรอนให้สาร x ซึงจะมีสาร ั ่ ตัวรับและถ่ายทอดกันเป็ นทอด ๆ คือ Feridoxin Cytochrome-b6 Cytochrome-f และ Plastocyanin แล้วกลับไปที่ P 700 การถ่ายทอดอิเลคตรอนเป็ นวัฎจักรพลังงานที่ ปลดปล่อยจากอิเลคตรอนนํ าไปสร้าง ATP ได้ 1 โมเลกุล และไม่มการสร้าง NADPH ี ้ เนื่องจากไม่มการแตกตัวของนํ้าจึงไม่ม ี O2 เกิดขึนด้วย ี การถ่ายทอดอิเลคตรอนเป็ นวัฎจักรเกิดจาก ระบบแสง II ไม่ทํางาน หรือขาด ประสิทธิภาพ ซึ่งอาจเกิดจากการถูกยับยังด้วยสารเคมี หรือได้รบแสงช่วงคลื่นมากกว่า ้ ั 690 นาโนเมตร การยับยังระบบแสง II ส่งผลให้ประสิทธิภาพการสังเคราะห์ดวยแสงลดลง ้ ้ + ิิ ่ ่ี ิ ด้วย NADP จากปฏิกรยาทีไม่ใช้แสงลดลงไปด้วย จึงเป็ นสาเหตุททําให้อเลคตรอนบาง ส่วนจากระบบแสง I กลับมาที่ Cytochrome f และ ระบบแสง I ตามลําดับ รูปที่ 10.2.7 การถ่ายทอดอิเลคตรอนเป็ นวัฎจักร (จาก George B. Johnson, 1995. The Living World หน้า 96) 2. การถ่ายทอดอิเลคตรอนไม่เป็ นวัฎจักร (Non Cyclic Electron Transfer) เป็ น การถ่ายทอดอิเลคตรอนผ่านทัง PS-I และ PS-II และมีการแตกตัวของนํ้าในปฏิกรยา ้ ิิ Phololysis โดยเริมจากสารสีใน PS-I และ PS-II ได้รบการกระตุนจากพลังงานแสง 2 ่ ั ้ โฟตอนพร้อมกัน P 700 ใน PS-I จะถ่ายทอดอิเลคตรอนให้สาร x (Ferridoxin Reducing Substant) และถ่ายทอดให้กบ Ferridoxin และ NADP+ ทํา P 700 ขาดอิเลคตรอนไป ั การทีจะคงสภาพเป็ นโมเลกุลทีให้อเลคตรอนของ P 700 ไว้ จึงต้องรับอิเลคตรอนจากสาร ่ ่ ิ อื่นทีมค่าความต่างศักย์ไฟฟ้าทีต่ํากว่า สารดังกล่าวนี้คอ Plastocyanin (Pc) ขณะเดียวกัน ่ ี ่ ื 436 CU 474
25.
Pc ก็จะรับอิเลคตรอนจาก PS-II
มาทดแทนโดย P 680 ทีได้รบการกระตุนจากพลังงาน ่ ั ้ แสง 2 โฟตอน ถ่ายทอด 2 อิเลคตรอนให้กบสาร Q (Plastoquinone Reducing Substant) ั และส่งต่อให้กบตัวรับอื่นตามลําดับ คือ Plastoquinone (Pq) Cytochrome b559 ั Cytochrome-f และ Plastocyanin (Pc) PS-II เมื่อสูญเสียอิเลคตรอนไปแล้วจะคงสภาพของโมเลกุล ผูให้อเลคตรอนโดย ้ ิ รับอิเลคตรอนจากการแตกตัวของนํ้าใน PS-II ในปฏิกรยา Photolysis ทีม ี Mn2- และ Clิิ ่ เป็ นตัวกระตุนโดยตรงและมีแสงเป็ นตัวช่วยกระตุนทางอ้อม ดังสมการ ้ ้ H2O Light 2H+ + 2e + ½ O 2 Mn2- , Cl- 2H+ จะส่งผ่านไปยัง Pq และเข้ารวมกับ NADP+ พร้อมกับรับอิเลคตรอนมาจาก PS-I ได้เป็ น NADPH + H+ ดังสมการ NADP+ + 2H+ + 2e- NADPH + H+ 2 อิเลคตรอน จะไปทดแทนอิเลคตรอนใน PS-I โดยถ่ายทอดไปทาง PS-II ซึงมีการ ่ ปลดปล่อยพลังงานทีนําไปสร้าง ATP ได้ 2 โมเลกุล ่ รูปที่ 10.2.8 การถ่ายทอดอิเลคตรอนไม่เป็ นวัฎจักรในการสร้าง ATP (ก) และการถ่ายทอดอิเลคตรอนผ่านสารทีมโปรตีนเป็ นองค์ประกอบในการสร้าง ATP (ข) ่ ี (จาก George B. Johnson, 1995. The Living World หน้า 96, 97) CU 474 437
26.
2. ปฏิกรยาทีไม่ตองใช้แสง (Light-Independent
Reaction) เป็ นปฏิกรยาตรึง CO2 ิิ ่ ้ ิิ (CO2 Fixation) สร้างสารประกอบคาร์โบไฮเดรต โดยใช้ ATP และ NADPH เป็ นแหล่ง พลังงาน ซึงกระบวนการเกิดเป็ นวงจรเรียกว่า วัฎจักรคัลวิน (Calvin Cycle) ตามชือของผู้ ่ ่ ค้นพบ คือ M. Calvin Calvin ทําการทดลองโดยใช้สาหร่าย Chlorella กับ 14CO2 ในระยะเวลาต่างกัน และหยุดปฏิกรยาในเวลาทีตองการ เพือนําสารประกอบทีเกิดขึนไปตรวจหา 14C โดยวิธี ิิ ่ ้ ่ ่ ้ Chromatography และ Radiogram ิิ ้ ผลการทดลอง ตรวจพบสารประกอบของ 14C ในปฏิกรยาการสังเคราะห์ดวยแสง ของสาหร่า ย ดังนี้ ในเวลา 7 วิน าที พบสาร 12 ชนิด เวลา 60 นาที พบมากชนิด ขึ้น ซึงรวมถึงกรดอินทรียและกรดอะมิโน ทีเวลา 5 วินาที พบ Phosphoglyceric Acid (PGA) ่ ์ ่ ซึงเป็ นสารประกอบคาร์บอน 3 อะตอม (3C) แต่มเี พียงอะตอมเดียวทีเป็ น 14C จึงน่าเชื่อ ่ ่ ว่าคาร์บอนอีก 2 อะตอมมาจากสารประกอบชนิดอื่นทีเข้ารวมกับ 14CO2 และไม่พบว่าใน ่ เซลล์ของสาหร่ายมีสารประกอบทีมคาร์บอน 2 อะตอม ่ ี 14 ิิ เมือให้ CO2 กับสาหร่ายไประยะหนึ่งแล้วหยุดปฏิกรยา ตรวจพบสาร Riburose ่ ่ 1, 5 Bisphosphate (RuBP) มี 14C เป็ นองค์ประกอบในจํานวนมากกว่าปกติ จึงเชือว่า 14 ่ ่ ้ RuBP เป็ นสารทีเข้ารวมกับ CO2 และพบว่าเมือ RuBP เพิมมากขึน PGA กลับลดลง ่ และในสภาวะทีเหมาะสมกับการสังเคราะห์ดวยแสง คือ มีแสง และ CO2 ปริมาณ PGA ่ ้ และ RuBP จะคงที่ และแสงไม่มผลต่อการสร้าง PGA เมือมี RuBP มากพอ แต่แสงทําให้ ี ่ RuBP เปลียนเป็ น PGA ได้ จึงเชือว่า RuBP เป็ นสารทีรวมกับ CO2 ได้เป็ น PGA และ ่ ่ ่ PGA ก็สามารถเปลียน RuBP ได้ ่ วัฎจักรคัลวิ น มี 3 ขัน ได้แก่ ้ 1. Carboxylation เป็ นการตรึง CO2 ด้วย RuBP เป็ นสารประกอบคาร์บอน 6 อะตอม โดยเริมจาก RuBP ซึ่งเป็ นนํ้ าตาลที่มคาร์บอน 5 อะตอมจับกับฟอสเฟต 2 หมู่ ่ ี ่ ั ่ เข้ารวมกับ CO2 ได้สารประกอบคาร์บอน 6 อะตอมทีไม่อยู่ตว เมือถูก Hydrolysis ด้วยนํ้า จะเปลียนเป็ นสารประกอบคาร์บอน 3 อะตอม 2 โมเลกุล คือ Phosphoglycerate, (PGA) ่ ่ เป็ นสารอินทรีย์ท่อยู่ตวชนิดแรก การตรึง CO2 เริมจาก RuBP 6 โมเลกุลรวมกับ CO2 ี ั 6 โมเลกุล ดังนันจึงได้ PGA 12 โมเลกุล ้ 6 RuBP + 6 CO2 438 12PGA CU 474
27.
2. Reduction เป็
นปฏิกรยาทีมการรีดวส์ PGA ไปเป็ นสารประกอบคาร์โบไฮเดรต ิิ ่ ี ิ ที่ป ระกอบด้ว ย คาร์บ อน 3 อะตอมกับ 1 หมู่ ฟ อสเฟต คือ Phosphoglyceraldehyde (PGAL) โดยอาศัยพลังงานจาก ATP และ NADPH ทีสร้างมาจากปฏิกรยาทีตองใช้แสง ่ ิิ ่ ้ 12PGA+12ATP+12NADPH+H+ 12PGAL+12ADP+12Pi+12NADP++12H+ 3. Regeneration and Synthesis เป็ นขันที่ PGAL เปลียนแปลงไป 2 ทาง ้ ่ ่ 3.1 สร้างสารประกอบทีใช้ตรึง CO2 โดย PGAL 10 โมเลกุล เปลียนกลับไปเป็ น ่ RuBP 6 โมเลกุลใหม่ โดยใช้พลังงานจาก ATP 6 โมเลกุล ซึงสลายให้ 6 ADP กับ 6 Pi ่ หมูฟอสเฟต 2 หมู่ ถูกนําไปใช้สร้าง RuBP จึงเหลือ 4 Pi ่ 10PGAL + 6ATP 6 RuBP + 6ADP + 4Pi กระบวนการในขันนี้ทาให้วฎจักรคัลวินดําเนินต่อไปได้อย่างต่อเนื่อง ้ ํ ั 3.2 สร้างสารอินทรียชนิดต่าง ๆ โดย PGAL 2 โมเลกุล อาจเปลียนเป็ นนํ้าตาล ์ ่ กลูโคส และแป้ง เพือนําไปใช้ในกระบวนการเมแทบอลิซมหรือเก็บสะสม ่ ึ รูปที่ 10.2.9 วัฎจักรคัลวินทีประกอบด้วย 3 ขันตอนสําคัญ ่ ้ th (จาก Purves et al. , 2003. Life : The Science of Biology, 7 Edition ในโครงการตําราวิทยาศาสตร์และ คณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 2 หน้า 77) CU 474 439
28.
การตรึงคาร์บอนไดออกไซด์ของพืช C3, C4
และ CAM การตรึง CO2 ในวัฎจักรคัลวินเกิดสารอินทรีย์ท่อยู่ตวชนิดแรก คือ PGA ซึ่ง ี ั ี ประกอบด้วย คาร์บอน 3 อะตอม กับ 1 หมู่ฟอสเฟต จึงเรียกการตรึง CO2 โดยวิธน้ีว่า ิิ วิถี C3 (C3- pathway) และเรียกพืชพวกนี้ว่า พืช C3 (C3- plant) ในปฏิกรยาต้องใช้ เอนไซม์ Ribulose Bisphosphate Carboxylase/Oxygenase (Rubisco) เป็ นสารเร่ง ปฏิกิรยา เอนไซม์น้ีนอกจากเร่งปฏิกิรยาการตรึง CO2 ด้วย RuBP แล้วยังสามารถเร่ง ิ ิ ้ ปฏิกรยาการตรึง O2 ด้วย RuBP ได้ดวย ทําให้การตรึง CO2 ของพืช C3 หลายชนิดลดลง ิิ ้ เนื่องจากมี O2 แข่งขันกับ CO2 ในการเข้ารวมกับ RuBP อัตราการสังเคราะห์ดวยแสงของ ่ ่ พืชจึงลดลง พืช C3 ได้แก่ ข้าวเจ้า ข้าวบาร์เลย์ ถัวและพืชทัวไป พืชบางชนิดมีกลไกเพิมความเข้มข้นของ CO2 ในสภาพแวดล้อมได้ดวย 2 กลไก ่ ้ สําคัญ ได้แก่ 1. กลไกการตรึงคาร์บอนด้วยการสังเคราะห์ดวยแสงแบบ C4 (C4 Photosynthetic ้ ่ Cabon Fixation) ซึงพบในพืชเขตร้อนหลายชนิด ได้แก่ อ้อย ข้าวโพด ข้าวฟาง และพืช ่ ตระกูลหญ้า จากการศึกษาของ Hatch และ Slack พบว่า พืชในกลุ่มดังกล่าวสามารถทําการ สังเคราะห์ดวยแสงได้ดี แม้จะมี CO2 น้อยมาก เขาคิดว่านอกจาก RuBP แล้ว น่ าจะมีสาร ้ ื ั ้ ี ชนิดอื่นทีใช้ตรึง CO2 ได้ จึงทําให้พชเหล่านี้สงเคราะห์ดวยแสงได้ดกว่าพืชชนิดอื่น ๆ จาก ่ ่ การทดลองจนสามารถสรุปขันการตรึง CO2 ทีเรียกว่า Hatch-Slack Pathway หรือ C4้ dicarboxylic acid pathway ซึงเกิดในเซลล์มโซฟิล (Mesophyll Cell) ดังนี้ ่ ี เริมจากการตรึง CO2 ด้วยสารประกอบคาร์บอน 3 อะตอมกับ 1 หมู่ฟอสเฟต คือ ่ Phosphoenolpyruvic Acid (PEP) ได้เป็ น Oxaloacetic Acid (OAA) ซึงมีคาร์บอน 4 ่ อะตอม และเปลียนต่อไปเป็ น Malic Acid หรือ Aspartic Acid ขึนอยู่กบชนิดของพืช โดย ่ ้ ั + ใช้ NADPH + H Malic Acid + NADP+ OAA + NADPH + H+ Malic Acid จะเปลียนเป็ น Pyruvic Acid และ CO2 ่ Malic Acid + NADP+ 440 Pyruvic Acid + CO2 + NADPH + H+ CU 474
29.
Pyruvic Acid จะเปลียนเป็
น PEP โดยใช้ ATP ส่วน CO2 จะเข้ารวมกับ RuBP ใน ่ Chloroplast ของเซลล์บนเดิลชีท (Bundle Sheath Cell) ในวัฎจักรคัลวินต่อไป ั เนื่องจากสารชนิดแรกทีเกิดจากการตรึง CO2 ด้วย PEP เป็ นสารประกอบคาร์บอน ่ ี ่ 4 อะตอม คือ OAA จึงนิยมเรียกการตรึง CO2 วิธน้ีวา C4 - pathway และเรียกพืชพวกนี้ ว่า พืช C4 (C4 - plant) รูปที่ 10.2.10 Hatch - Slack Pathway ของพืช C4 (จาก George B. Johnson, 1995. The Living World หน้า 99) ข้อแตกต่างระหว่างพืช C3 และ C4 ี ่ 1. พืช C4 สามารถตรึง CO2 ได้ดกว่าพืช C3 มาก โดยเฉพาะในสภาพแวดล้อมทีม ี ่ ั อุณหภูมสงจะมี CO2 ตํ่า พืช C4 จะเพิมความเข้มข้นของ CO2 ในเซลล์บนเดิลชีทได้ ิู ้ 2. พืช C4 สามารถสังเคราะห์ดวยแสงในสภาพความเข้มแสงมาก ๆ ได้ ในขณะที่ ้ ุ่ ่ ่ํ พืช C3 สังเคราะห์ดวยแสงได้มากทีสดทีระดับความเข้มแสงทีจากัด CU 474 441
30.
3. พืช C4
สังเคราะห์ดวยแสงทีอุณหภูมพอเหมาะ (Optimum) ค่อนข้างสูง คือ ้ ่ ิ ่ ื ่ ่ี ิ 30-40 C ในขณะทีพช C3 อยูทระดับอุณหภูมประมาณ 25 °C ่ ่ ้ ํ 4. ปริมาณ O2 ทีเพิมขึน (3-20 %) ไม่ทาให้ประสิทธิภาพการสังเคราะห์แสงของ ี พืช C4 ลดลง แต่มผลในพืช C3 5. ในสภาพทีมแสงจ้าและอุณหภูมสงจะมีผลต่อการหายใจด้วยแสง (Photorespiration) ่ ี ิู ี ่ ้ ในพืช C3 ทําให้มการสลายคาร์โบไฮเดรต ซึงเป็ นผลผลิตจากกระบวนการสังเคราะห์ดวย แสง กรณีเช่นนี้จะไม่เกิดขึนในพืช C4 ้ 2. กลไก Crassulacean Acid Metabolism (CAM) Pathway พบในพืชทะเลทราย ตระกูล Crassulaceae เช่น กุหลาบหินและพืชในสกุลอื่น ๆ เช่น กระบองเพชร สับปะรด วานิลา และอากาเว่ เป็ นต้น พืชทะเลทรายมีการปรับตัวโดยการลดรูปของปากใบ เพื่อป้องกันการสูญเสียนํ้ า ่ ่ และปากใบจะปิดในเวลากลางวัน เปิดเวลากลางคืน CO2 จะเข้าสูใบทางปากใบไปทีเซลล์ม ี โซฟิล มีการตรึง CO2 ด้วย Phosphoenolpyruvic Acid (PEP) ได้ Oxaloacetic Acid (OAA) และเปลียนไปเป็ นกรดมาลิก (Malic Acid) ไปเก็บสะสมไว้ในแวคิวโอล เมือเริมมี ่ ่ ่ แสงในเวลากลางวันปากใบจะปิ ดและมีการเคลื่อนย้ายกรดมาลิกออกจากแวคิว-โอล เพื่อ ่ ทําปฏิกรยาปล่อย CO2 ออกจากกรดมาลิก (Decarboxylation) เข้าสูกระบวนการตรึง CO2 ิิ ในวัฎจักรคัลวินตามปกติ จะเห็นได้ว่ากระบวนการ CAM - pathway พืชจะปิ ดปากใบในเวลากลางวัน ทําให้มความเข้มข้นของ CO2 สูงมาก อัตราการหายใจด้วยแสงจึงลดลงตํ่ามาก และการ ี ตรึง CO2 ด้วย PEP เกิดในเวลากลางคืน เนื่องจากในเวลากลางวันมีแสง กรดมาลิกที่ ปล่อยออกจากแวคิวโอลจะยับยังการทํางานของเอนไซม์ทกระตุนการตรึง CO2 ด้วย PEP ้ ่ี ้ ส่วนในเวลากลางคืนไม่มแสง กรดมาลิกจะไม่ยบยังการทํางานของเอนไซม์ทกระตุนการ ี ั ้ ่ี ้ ตรึง CO2 ด้วย PEP จึงทํางานได้ตามปกติ ้ เนื่องจากการตรึง CO2 ด้วย PEP ของพืชบริเวณแห้งแล้ง หรือทะเลทราย พบครัง แรกในพืชตระกูล Crassulaceae จึงเรียกพืชพวกนี้วา Crassulacean Acid Metabolism ่ Plant หรือ พืช CAM ° 442 CU 474
31.
รูปที่ 10.2.11 การตรึง
CO2 ด้วย PEP ของพืช CAM (จาก โครงการตําราวิทยาศาสตร์และคณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 2 หน้า 93) การหายใจด้วยแสงของพืช (Photorespiration) การหายใจด้วยแสงเป็ นวัฎจักรการตรึง O2 ด้วย RuBP และมีการปล่อย CO2 เช่นเดียวกับการหายใจตามปกติของพืช แต่การหายใจด้วยแสงทําให้พชสูญเสียคาร์บอนที่ ื ตรึงไว้ โดยไม่มการสร้างพลังงานทีพชจะนําไปใช้ได้เพราะเป็ นกระบวนการทีเกิดพร้อมกับ ี ่ ื ่ การตรึง CO2 ในวัฎจักรคัลวิน การหายใจด้ว ยแสงมีก ารเปลี่ย นแปลงของสารเป็ น วงจรต่ อ เนื่ อ งกัน เรีย กว่ า วัฎจักรกระบวนการหายใจด้วยแสง (Photorespiratory Carbon Oxidation Cycle ; PCO ิิ Cycle) เริมจากการตรึง O2 ด้วย RuBP โดยมีเอนไซม์ Rubisco เป็ นตัวเร่งปฏิกรยา ่ สร้าง 2 - phosphoglycolate ซึงจะถูก Hydrolysis ไปเป็ น Glycolate และออกจากคลอโร่ พลาสต์เข้าสู่ Peroxisome ทีน่ี Glycolate จะเกิด 2 ปฏิกรยาเปลียนไปเป็ น Glycine เพือ ่ ิิ ่ ่ เข้าสู่ Mitochondria โดย 2 โมเลกุลของ Glycine จะเปลียนเป็ น Serine และ CO2 ซึงเป็ น ่ ่ ปฏิกรยาทีมการสูญเสียคาร์บอนไป Serine จะกลับเข้าสู่ Peroxisome เพื่อเปลียนเป็ น ิิ ่ ี ่ Glycerate กลับเข้าสูคลอโรพลาสต์ และเปลียนเป็ น Phosphoglycarate (PGA) 3 โมเลกุล ่ ่ ในวัฎจักรคัลวิน วัฎจักรกระบวนการหายใจด้วยแสง (PCO Cycle) เป็ นวัฎจักรทีทาให้พชสามารถ ่ ํ ื นํ าคาร์บอนจากโมเลกุลคาร์โบไฮเดรตที่ถูกออกซิไดซ์กลับมาใช้ได้ใหม่ โดยสร้างเป็ น PGA (3C) 1 โมเลกุล จาก Phosphoglycolate (2C) 2 โมเลกุล และสูญเสียสารประกอบ ่ คาร์บอนในโมเลกุล CO2 1 โมเลกุล ซึงสรุปได้ว่าในวัฎจักร PCO นําคาร์บอนกลับคืนมา CU 474 443
32.
ได้ถง 75 %
(3 อะตอมของ PGA 1 โมเลกุล จาก 4 อะตอม ของ Phosphoglycolate 2 ึ โมเลกุล) โดยวัฎจักร PCO ต้องมีวฎจักรคัลวินเป็ นตัวนํา PGA ไปสร้าง RuBP ั ในคลอโรพลาสต์ แม้วา O2 และ CO2 ในรูปของสารละลายจะแข่งขันกันในการเข้า ่ จับกับเอนไซม์ Rubisco ในการทําปฏิกรยากับ RuBP และสารละลายทีมความเข้มข้นของ ิิ ่ ี O2 และ CO2 เท่ากัน Rubisco จะจับกับ CO2 ได้เร็วกว่าถึง 80 เท่าก็ตาม แต่เนื่องจาก สารละลายในสภาวะสมดุลกับอากาศปกติทอุณหภูม ิ 25 °C จะมี O2 มากกว่า คือ 24 เท่า ่ี ้ จึงทําให้อตราการตรึง CO2 มากกว่าอัตราการตรึงของ O2 ประมาณ 3 เท่า เท่านัน ั สรุปได้ว่าวัฎจักรคัลวินและวัฎจักร PCO มีความสําคัญในการรักษาสมดุลของ ี คาร์บอนในพืช โดยวัฎจักรคัลวินมีการตรึง CO2 และปล่อย O2 พืชมีชวะมวล (Biomass) ี เพิมขึน ในขณะทีวฎจักร PCO มีการตรึง O2 และปล่อย CO2 พืชมีชวะมวลลดลง วัฎจักร ่ ้ ่ั ทัง 2 เกิดขึนในเซลล์เดียวกัน ในอัตราการตรึง CO2 3 ส่วน และการตรึง O2 1 ส่วน ทําให้ ้ ้ การตรึงคาร์บอนกับการสูญเสียคาร์บอนมีความสมดุล รูปที่ 10.2.12 สมดุลของคาร์บอนในวัฎจักรคัลวินและวัฎจักร PCO ของพืช C3 (จาก โครงการตําราวิทยาศาสตร์และคณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 2 หน้า 85) ปัจจัยที่มีผลต่ออัตราการสังเคราะห์ด้วยแสง 1. แสงสว่าง (Light) อัตราการสังเคราะห์ดวยแสงของพืชขึนอยู่กบความเข้มของ ้ ้ ั แสง ช่วงคลื่นแสง และระยะเวลาการได้รบแสง สภาพความเข้มของแสงน้ อย แสงเป็ น ั ั ป จ จัย จํา กัด ของการสังเคราะห์ด้ว ยแสง การเพิ่ม ความเข้ม ของแสงจึง ทํา ให้อ ต ราการ ั สังเคราะห์ดวยแสงเพิมขึน แต่ถาพืชได้รบแสงทีมความเข้มมากเป็ นเวลานานเกินไป จะทํา ้ ่ ้ ้ ั ่ ี ให้ก ารสัง เคราะห์ด้ว ยแสงหยุ ด ลงได้ เนื่ อ งจากคลอโรฟิ ล ถู ก กระตุ้น มากเกิน ไป และ ออกซิเจนทีเกิดขึนถูกใช้ไปในกระบวนการหายใจด้วยแสง (Photorespiration) ซึงมีการ ่ ้ ่ 444 CU 474
33.
สลายโมเลกุลของคาร์โบไฮเดรต ทําให้สญเสียคาร์บอน อัตราการสังเคราะห์ดวยแสงจึงไม่ ู ้ ตอบสนองต่อแสง ั ่
ี 2. CO2 ความเข้มข้นของ CO2 ไม่เป็ นปจจัยจํากัดในเวลาทีมแสงมาก ในสภาวะที่ ้ มี CO2 แต่แสงน้อย อัตราการสังเคราะห์ดวยแสงจะน้อย และทําให้ CO2 มากในเวลานาน เกินไป อัตราการสังเคราะห์ดวยแสงจะสูงในตอนต้นของปฏิกรยาต่อจากนันจะลดลง ้ ิิ ้ รูปที่ 10.2.13 ความเข้มของแสงและปริมาณ CO2 ทีมอทธิพลต่อการสังเคราะห์ดวยแสง ่ ีิ ้ 3. อุณหภูม ิ (Temperature) พืชจะมีการสังเคราะห์ดวยแสงได้ดท่อุณหภูม ิ ้ ี ี ° ิู พอเหมาะ (Optimum Temperature) คือ 30-35 C ถ้าอุณหภูมสงเกินไป เช่น 60 °C เอนไซม์จะไม่ทํางาน เนื่องจากโปรตีนเสียสภาพ ถ้าอุณหภูมต่ําเกินไป เอนไซม์จะทํางาน ิ ไม่ดีเ ท่า ที่ค วร การแพร่ข อง CO2 เข้า สู่ใ บเกิด ช้า มาก นอกจากนี้ ย ง มีผ ลทางอ้อ ม คือ ั อุณหภูมสงอัตราการหายใจเพิมสูงขึน อัตราการสังเคราะห์ดวยแสงจะลดลง เพราะมีการ ิู ่ ้ ้ สลายโมเลกุลคาร์โบไฮเดรตไปใช้ในการหายใจ รูปที่ 10.2.14 กราฟแสดงปริมาณ CO2 ในบรรยากาศปกติ และสูงกว่าปกติทอุณหภูมต่าง ๆ มีอทธิพลต่อการ ่ี ิ ิ สังเคราะห์ดวยแสงในพืชพวกมันฝรัง่ ้ CU 474 445
34.
4. เกลือแร่ ธาตุไนโตรเจน
และแมกนีเซียม ของเกลือในดิน มีความสําคัญต่ออัตรา การสังเคราะห์ด้วยแสง เนื่องจากเป็ นองค์ประกอบของคลอโรฟิ ล ถ้าขาดจะมีผลต่อการ สร้างคลอโรฟิล ธาตุเหล็กแม้ไม่ใช้องค์ประกอบของคลอโรฟิลโดยตรงแต่กมผลต่อการสร้าง ็ ี คลอโรฟิล ซึงขาดไม่ได้เช่นกัน ่ 5. โครงสร้างของใบ 1) ใบของพืช มีโครงสร้างที่เอื้อต่อการรับแสง และแลกเปลียนแก๊ส เริมตังแต่ ่ ่ ้ เซลล์ชนนอกสุด คือ เอพิเดอร์มส จะปล่อยให้แสงส่องผ่านไปในใบได้ โดยเฉพาะใบพืชพืน ั้ ิ ้ ป่าเขตร้อน เอพิเดอร์มสจะมีลกษณะคล้ายเลนส์นูน ซึงรวมแสงให้มความเข้ม แสงทีผ่าน ิ ั ่ ี ่ ไปทีคลอโรฟิลจึงมีความเข้มมากกว่าแสงนอกใบ ่ ชันมีโซฟิล มีเซลล์พาลิเสด ลักษณะเป็ นแท่งตัง จะดูดแสงส่วนหนึ่งโดยใช้สารสี ้ ้ ั ต่าง ๆ ในคลอโรพลาสต์ แสงส่วนทีเหลือจะผ่านเข้าสู่ช่องว่างในเซลล์สปนจี ซึ่งมีรูปร่าง ่ หลายแบบ ในช่องว่างที่มน้ํ าและอากาศจะช่วยให้แสงสะท้อนจากรอยต่อระหว่างนํ้ ากับ ี ั อากาศไปหลายทิศทาง โอกาสการดูดแสงของเซลล์สปนจีมเี พิมขึน ่ ้ พืชทีอยู่บริเวณแสงมากจนอาจเป็ นอันตราย จะมีโครงสร้างพิเศษ เพื่อป้องกัน ่ การได้รบแสงมาก เช่น มีขน มีชนไขมันเคลือบ ช่วยในการสะท้อนกลับของแสง เป็ นต้น ั ั้ 2) ใบพืชควบคุม การรับแสงโดยการเคลื่อนทีของคลอโรพลาสต์ในเซลล์ และ ่ การเคลื่อนไหวของใบ พืชพวกถัวและฝ้าย สามารถเคลื่อนใบตามแสงอาทิตย์ทําให้ได้รบ ่ ั แสงมาก โดยเฉพาะช่วงเวลาเช้าและเย็น ซึ่งเป็ นช่วงทีอากาศมีความชืนและอุณหภูมต่ํา ่ ้ ิ กว่าตอนเทียงวัน ในขณะทีพชบางชนิดมีการเคลื่อนใบหลบหลีกแสงอาทิตย์ เพื่อลดการ ่ ่ ื ได้รบแสงมาก ซึงมักเป็ นกับพืชบริเวณขาดนํ้า ั ่ 3) พืชมีการปรับตัว ทังเซลล์ใบและลําต้น โดยพืชทีอยู่ในทีมแสงน้อย มักมีใบ ้ ่ ่ ี หนาเพือเพิมชันเซลล์ ่ ่ ้ 4) การจัดโครงสร้างของเรือนพุ่ม โดยจัดเรียงตัวของใบ ลําต้น กิง และก้านใบ ่ ให้สามารถแข่งขันเพื่อรับแสง เช่น ใบมะละกอ มีกานใบยาว และจัดเรียงตัวรอบลําต้น จึง ้ ทําให้ใบทุกใบได้รบแสงเต็มที่ ั 446 CU 474
35.
10.3 การสืบพันธุและการเจริญเติบโตของพืชดอก ์ (Flowering Plant
Reproduction and Development) การสืบพันธุ์แบบอาศัยเพศของพืชดอก มีดอก (Flower) เป็ นอวัยวะทีทําหน้าที่ ่ สืบพันธุ์ โดยทําหน้าทีสร้างเซลล์สบพันธุ์ คือ ละอองเรณู (Microspore) ซึ่งเป็ นเซลล์ ่ ื สืบพันธุเพศผูและไข่ (Egg) ซึงเป็ นเซลล์สบพันธุเพศเมียขึนมาผสมพันธุกนเป็ นเซลล์ ไซ ์ ้ ่ ื ์ ้ ์ ั โกต (Zygote) และเพิมจํานวนโดยการแบ่งเซลล์ (Cell Division) ทีจะเติบโตและ ่ ่ เปลียนแปลงเซลล์ไปทําหน้าทีเฉพาะ (Cell Growth and Differentiation) อย่างเป็ น ่ ่ ขันตอน สร้างรูปร่าง (Form) ทีเหมาะสมกับหน้าที่ (Function) เฉพาะของต้นพืชทีเรียกว่า ้ ่ ่ การเจริญ (Development) ดอกและส่วนประกอบของดอก ดอก (Flower) คือ ส่วนทีเปลียนแปลงมาจากกิงและใบเพื่อทําหน้าทีสบพันธุและ ่ ่ ่ ่ ื ์ แพร่พนธุ์ เพื่อการดํารงพันธุ์มให้สูญสินไปจากโลก กิงทีเปลี่ยนมาเป็ นดอกจะมีปล้องสัน ั ิ ้ ่ ่ ้ มาก ซึงแตกต่างจากกิงทัวไปถือว่าเป็ นกิงพิเศษ เป็ นกิงทีการเจริญเติบโตมีขอบเขตจํากัด ่ ่ ่ ่ ่ ่ คือ จะไม่มการเจริญทีสวนปลาย (Apical Growth) ต่อไปอีก ส่วนประกอบทีสาคัญของ ี ่่ ่ํ ดอก ได้แก่ กลีบเลียง (Sepal) กลีบดอก (Petal) เกสรตัวผู้ (Stamen) และเกสรตัวเมีย ้ (Pistil) ทังหมดนี้เป็ นส่วนทีเปลียนแปลงมาจากใบ โดยกลีบเลี้ยงและกลีบดอกจัดเป็ น ้ ่ ่ อวัยวะช่วยสืบพันธุ์ (Accessory Organs) ส่วนเกสรตัวผูและเกสรตัวเมีย จัดเป็ นอวัยวะ ้ สืบพันธุ์ (Reproductive Organs) ตา (Bud) ทีให้กําเนิดดอกอาจเป็ นตาดอก (Flower Bud) ซึงจะเป็ นดอกอย่างเดียว ่ ่ หรือตาผสม (Mixed Bud) คือ ตาทีเป็ นทังกิงใบและดอกร่วมกัน เกิดได้ทงทีปลายยอดหรือ ่ ้ ่ ั้ ่ ั ปลายกิงของลําต้น การพัฒนาตาไปเป็ นดอกของพืชต้องอาศัยปจจัยต่าง ๆ ทังภายนอก ่ ้ ั และภายในของพืช ปจจัยภายนอก ได้แก่ แสงสว่าง อุณหภูม ิ นํ้า แร่ธาตุ และสารอาหารที่ พืชสร้างขึน ปจจัยภายใน ได้แก่ อายุ พันธุกรรม และฮอร์โมนพืช ้ ั CU 474 447
36.
โครงสร้างของดอก (Structure of
Flower) ดอกทีมสวนประกอบครบ (Complete Flower) จะจัดเรียงตัวเป็ นชันหรือวงติดกัน ่ ี่ ้ 4 ชัน จากชันนอกเข้าไป ดังนี้ ้ ้ ก. ข. รูปที่ 10.3.1 ดอกไม้แสดงส่วนประกอบของดอกครบ (ก) และแผนภาพแสดงชันต่าง ๆ ของดอก (ข) ้ 1. ชันกลีบเลียง (Calyx) ประกอบด้วย กลีบเลียง (Sepals) ดอกไม้ทวไป กลีบเลียงมี ้ ้ ้ ั่ ้ สีเขียว แต่อาจมีสอ่นได้ มีผวเรียบ บางชนิดมีหนาม และขน ทําหน้าทีห่อหุมส่วนทีอยู่ขาง ีื ิ ่ ้ ่ ้ ในไว้ทงหมด จึงช่วยป้องกันอันตราย ได้แก่ อุณหภูม ิ ความชืน รวมถึงแมลงและศัตรูอ่น ๆ ั้ ้ ื พืชแต่ละชนิดมีกลีบเลี้ยงจํานวนไม่เท่ากัน ดอกไม้บางอย่างมีกลีบเลี้ยงติดกันเป็ นหลอด หุมดอก (Synsepalous Flower) บางชนิดมีกลีบเลียงแยกกัน (Polysepalous Flower) ้ ้ ดอกบางชนิดมีกลีบสีเขียวเป็ นเส้นเล็ก ๆ ทีใต้กลีบเลียง เรียกว่า ริวประดับ (Epicalyx) ่ ้ ้ 2. ชันกลีบดอก (Corolla) ประกอบด้วย กลีบดอก (Petals) สีต่าง ๆ ซึงเกิดจาก ้ ่ สารสีชนิดต่าง ๆ ทีอยู่ในเซลล์ บางชนิดมีกลินหอมทีเกิดจากนํ้ามันหอมระเหย (Essential ่ ่ ่ Oil) ทีกลีบดอกสร้างขึน บางชนิดมีต่อมนํ้ าหวาน (Nectary Gland) ทีโคนกลีบสร้าง ่ ้ ่ นํ้าหวาน (Nectar) ได้ ทังสี กลิน และนํ้าหวาน เป็ นสิงทีชวยล่อแมลงให้เข้ามาถ่ายละออง ้ ่ ่ ่ ่ เรณู (Pollination) พืชแต่ละชนิดมีกลีบดอกจํานวนไม่เท่ากัน แต่มกจะเท่ากับหรือทวีคูณ ั ของกลีบเลียง กลีบดอกอาจมีโคนกลีบเชือมติดกันหรือติดกันตลอดทังกลีบ (Synpetalous ้ ่ ้ Flower) หรือแยกกันเป็ นกลีบ ๆ หรือแยกเฉพาะปลายกลีบ (Polypetalous Flower) 3. ชันเกสรตัวผู้ (Androecium) ประกอบด้วย เกสรตัวผู้ (Stamen) ทําหน้าทีสร้าง ้ ่ เซลล์สบพันธุ์เพศผู้ เกสรตัวผูมจํานวนแตกต่างกันในพืชแต่ละชนิด พวกทีมหลายอันอาจ ื ้ ี ่ ี ติดกันหรือแยกกัน และอาจมีสวนทีไปเชือมติดกับส่วนอื่นของดอกก็ได้ เกสรตัวผูแต่ละอัน ่ ่ ่ ้ ประกอบด้วย ก้านเกสรตัวผู้ (Filament) มีลกษณะเป็ นเส้นเล็กยาว ตอนปลายมีอบเรณู ั ั 448 CU 474
37.
(Anther) ลักษณะเป็ นทรงกระบอก
กลมรี แบ่งเป็ น 2 พูตดกัน ภายในแต่ละพู ประกอบด้วย ถุง ิ เรณู (Pollen Sac) 2 ถุง อับเรณู 1 อัน จึงมีถุงเรณู 4 ถุง ภายในถุงมี Microspore Mother Cell ทีจะทําหน้าทีสร้างละอองเรณู (Microspore หรือ Pollen Grain) ซึงมีลกษณะเป็ นเม็ด ่ ่ ่ ั เล็ก ๆ จํานวนมาก ละอองเรณูทําหน้าทีเป็ นเซลล์สบพันธุเพศผู้ (Male Gametes) เมื่อ ่ ื ์ ละอองเรณูแก่ อับเรณูจะแตก ละอองเรณูจะปลิวออกมา และอาจตกลงบนยอดเกสรตัวเมีย ทําให้เกิดการผสมพันธุได้ ์ 4. ชันเกสรตัวเมีย (Gynaecium) ประกอบด้วย เกสรตัวเมีย (Pistil or Carpel) ้ เป็ นส่วนที่อยู่ในสุดของดอก ทําหน้ าที่สร้างเซลล์สบพันธุ์เพศเมีย ส่วนล่างสุดของเกสร ื ตัวเมีย คือ รังไข่ (Ovary) มีลกษณะเป็ นกระเปาะพองโต ถัดขึนมาเป็ นก้านเกสรตัวเมีย ั ้ (Style) ซึ่งอาจจะยาวหรือสันขึนอยู่กบชนิดของพืชทีปลายสุดของก้านเกสรตัวเมีย คือ ้ ้ ั ่ ยอดเกสรตัวเมีย (Stigma) มักมีน้ําเหนียว ๆ สําหรับยึดติดกับละอองเรณู ดอกบางชนิด ไม่มกานเกสรตัวเมีย ยอดเกสรตัวเมียจึงติดอยู่บนรังไข่ (Sessile Stigma) พืชบางชนิดมี ี ้ เกสรตัวเมียหลายอัน เช่น จําปี จําปา กุหลาบ เป็ นต้น รังไข่ (Ovary) เป็ นส่วนที่สร้างเซลล์สบพันธุ์ของพืชโดยภายในรังไข่มโอวุล ื ี (Ovule) ซึงภายในมี Megaspore Mother Cell ทีจะทําหน้าทีสร้างเซลล์สบพันธุเพศเมีย ่ ่ ่ ื ์ คือ เซลล์ไข่ (Egg Cell) โอวุล เป็ นโครงสร้างทีประกอบด้วยผนังโอวุล (Integument) 1-2 ่ ชัน โดยมีรูเปิ ดเล็ก ๆ เรียกว่า ไมโครไพล์ (Micropyle) อยู่ด้านตรงข้ามกับก้านโอวุล ้ (Funiculus) ภายในมีกลุ่มเซลล์นิวเซลลัส (Nucellus) ซึ่งจะเจริญและพัฒนาเป็ น Megaspore Mother Cell ทีจะสร้างเซลล์ไข่ ต่อไป ่ ส่วนประกอบของดอกทัง 4 ชันติดตังบนฐานรองดอก (Receptacle) ซึงติดต่อกับ ้ ้ ้ ่ ก้านดอก (Pedicel หรือ Peduncle) การติดตังของรังไข่บนฐานรองดอกมีแบบต่าง ๆ ้ ได้แก่ 1. Superior Ovary คือ รังไข่ทอยู่บนฐานรองดอกในตําแหน่งทีสงกว่าส่วนประกอบ ่ี ู่ อื่น ๆ ของดอก คือ กลีบเลียง กลีบดอกและเกสรตัวผู้ ้ 2. Inferior Ovary คือ รังไข่ทอยู่บนฐานรองดอกในตําแหน่ งทีต่ํากว่ากลีบเลียง ่ี ่ ้ กลีบดอกโดยมีผนังรังไข่ เชือมติดกับส่วนประกอบของดอก ่ 3. Half Inferior หรือ Subinferior Ovary คือ รังไข่ทตดตังบนฐานรองดอกใน ่ี ิ ้ ลักษณะกิงใต้ชนกลีบเลียง กลีบดอก และเกสรตัวผู้ เนื่องจากลักษณะฐานรองดอกเป็ นรูป ่ ั้ ้ ถ้วย CU 474 449
38.
รูปที่ 10.3.2 รังไข่ชนิดต่าง
ๆ (จาก โครงการตําราวิทยาศาสตร์และคณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 1) ประเภทของดอก (Type of Flower) 1. จําแนกตามส่วนประกอบของดอก แบ่งได้เป็ น 2 ชนิด ได้แก่ ดอกสมบูรณ์ (Complete Flower) คือ ดอกทีมสวนประกอบครบทัง 4 ชัน ่ ี่ ้ ้ ดอกไม่สมบูรณ์ (Incomplete Flower) คือ ดอกทีมสวนประกอบไม่ครบทัง ่ ี่ ้ 4 ชัน ้ 2. จําแนกตามลักษณะของเพศ แบ่งได้เป็ น 2 ชนิด ได้แก่ ดอกสมบูรณ์เพศ (Perfect Flower) คือ ดอกทีมทงเกสรตัวผูและเกสรตัวเมียอยู่ใน ่ ี ั้ ้ ดอกเดียวกัน เช่น พูระหง กุหลาบ บัว เฟื่องฟ้า เป็ นต้น ่ ดอกไม่สมบูรณ์เพศ (Imperfect Flower) คือ ดอกทีมเี กสรตัวผูหรือเกสรตัวเมีย ่ ้ เพียงอย่างเดียว ดอกทีมเี กสรตัวผูเรียกว่า ดอกเพศผู้ (Staminate Flower) ดอกทีมเี กสร ่ ้ ่ ตัวเมีย เรียกว่า ดอกเพศเมีย (Pistillate Flower) พืชทีมดอกเพศผู้ และดอกเพศเมียอยู่ในต้นเดียวกัน เรียกว่า พืชแยกเพศร่วมต้น ่ ี (Monoecious Plant) เช่น หมาก มะพร้าว ข้าวโพด เป็ นต้น ส่วนพืชทีมดอกเพศผูและ ่ ี ้ ั ดอกเพศเมีย อยูกนคนละต้น เรียกว่า พืชแยกเพศต่างต้น (Dioecious Plant) เช่น ฟกทอง ่ ั 3. จําแนกตามจํานวนดอก แบ่งได้แป็ น 2 ชนิด ได้แก่ ดอกเดียว (Solitary Flower) คือ ดอกเดียวบนก้านดอกเดียว ่ ่ ดอกช่อ (Inflorescence Flower) คือ กลุ่มดอกบนก้านดอกเดียวกันแต่ละดอก ย่อย (Floret) จะมีกานดอกย่อย (Pedicel) ติดอยูบนก้านของช่อดอก ้ ่ 4. จําแนกตามการติดตังบนฐานรองดอก แบ่งได้เป็ น 3 ชนิด ได้แก่ ้ 450 CU 474
39.
Hypogenous Flower คือ
ดอกทีมฐานรองดอก นูนสูง เกสรตัวเมียจึงอยูสงกว่า ่ ี ่ ู ส่วนของกลีบเลียง กลีบดอกและเกสรตัวเมีย ดอกชนิดนี้จงมีรงไข่ชนิด Superior Ovary ้ ึ ั Epigynous Flower คือ ดอกทีสวนของกลีบเลียง กลีบดอก และเกสรตัวผูตดบน ่่ ้ ้ ิ ฐานรองดอกทีอยู่เหนือเกสรตัวเมีย เป็ นดอกทีมรงไข่ชนิด Inferior Ovary ได้แก่ แตงกวา ่ ่ ีั ั บวบ ฟกทอง ชมพู่ เป็ นต้น Perigynous Flower คือ ดอกทีมสวนของกลีบเลียง กลีบดอกและเกสรตัวผูอยู่ ่ ี่ ้ ้ ติดบนฐานรองดอกในระดับเดียวกับรังไข่ เพราะลักษณะที่โค้งรูปถ้วยของฐานรองดอก ดอกชนิดนี้รงไข่เป็ นได้ 2 แบบ คือ Half Superior Ovary และ Half Inferior Ovary ขึนอยู่ ั ้ กับลักษณะของฐานรองดอก รูปที่ 10.3.3 แผนภาพชนิดของดอกตามการติดตังบนฐานรองดอก Hypogenous Flower (ก) ้ Epigynous Flower (ข) Perigynous Flower (ค,ง และ จ) (จาก โครงการตําราวิทยาศาสตร์และคณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 1) การสร้างเซลล์สืบพันธุในพืชดอก ์ การสร้างเซลล์สืบพันธุเพศผูหรือไมโครสปอร์ (Microspore genesis) ์ ้ การสร้างเซลล์สบพันธุเพศผูของพืชดอก เกิดขึนภายใน Pollen Sac ของอับเรณู ื ์ ้ ้ (Anther) โดยกลุมเซลล์ Microspore Mother Cell ซึงเป็ นเซลล์เนื้อเยื่อเจริญ (Meristematic ่ ่ Cell) ทีม ี 2 n โครโมโซมจะแบ่งตัวแบบไมโอซิสได้ไมโครสปอร์ 4 เซลล์ (4 Microspore ่ Cell) ทีเป็ นเซลล์ n โครโมโซม แต่ละเซลล์จะแบ่งนิวเคลียสแบบไมโตซิส 1 ครัง ได้ 2 ่ ้ นิวเคลียสทีเป็ น n โครโมโซม นิวเคลียสอันหนึ่ง คือ Generative Nucleus อีกนิวเคลียส ่ คือ Tube Nucleus (ซึงต่อไป Generative Nucleus จะแบ่งไมโตซิสอีก 1 ครัง ได้เป็ น 2 ่ ้ Sperm Nucleus เพื่อทีจะไปผสมกับไข่และโพลาร์นิวเคลียส ต่อไป) ในขณะเดียวกัน ่ Microspore ก็สร้างผนังให้หนาขึน ผนังชันนอกอาจมีการพัฒนาไปเป็ นลักษณะต่าง เช่น ้ ้ CU 474 451
40.
เป็ นหนามเล็ก ๆ
Microspore ละอองเรณู (Pollen Gain) ระยะนี้จะมีรูปร่างต่าง ๆ ตามชนิดของพืชและเรียกว่า ก. ข. รูปที่ 10.3.4 การสร้าง Microspore ของอับเรณู (ก) การแบ่งไมโอซิสของ Microspore (Pollen) Mother Cell และไมโตซิสของ Microspore (ข) การสร้างเซลล์สืบพันธุเพศเมีย หรือเมกะสปอร์ (Megaspore genesis) ์ การสร้างเซลล์สบพันธุเพศเมีย คือ เซลล์ไข่ของพืชดอก เกิดในโอวุลซึงอยู่ในรังไข่ ื ์ ่ กลุมเซลล์ในโอวุลทีเรียกนิวเซลลัส (Nucellus) ทีอยูคอนไปทางด้านไมโครไพล์ มีเซลล์ขนาด ่ ่ ่ ่ ่ ใหญ่ คือ Megaspore Mother Cell (2n) แบ่งตัวแบบไมโอซิสได้ 4 เซลล์ คือ Megaspore Cell (n) เรียงตัวในแนวตัง ต่อมา 3 เซลล์ ทางไมโครไพล์ สลายตัวไป 1 Megaspore ที่ ้ เหลือเจริญขึนและแบ่งนิวเคลียสแบบไมโตซิส 3 ครัง ได้ 8 นิวเคลียสทีไม่มผนังหุม และ ้ ้ ่ ี ้ จัดแบ่งเป็ น 2 กลุ่ม ๆ ละ 4 นิวเคลียส แยกกันไปอยู่ทางด้านไมโครไพล์ กับด้านตรงข้าม ไมโครไพล์ คือ คาลาซา (Chalaza) ต่อมานิวเคลียสของแต่ละกลุ่มจะมารวมกันบริเวณ กลาง เกิดเป็ นนิวเคลียส 3 กลุ่ม ระยะนี้ Megaspore จะมีสภาพเป็ นถุงต้นอ่อน (Embryo Sac) กลุ่มนิวเคลียส 2 กลุ่มทีไมโครไพล์และคาลาซา จะมีเยื่อบาง ๆ แบ่งไซโตพลาสซึม ่ เกิดเป็ นเซลล์ คือ 3 Anti Podal Cell อยู่ทางด้านคาลาซา ด้านไมโครไพล์ ประกอบด้วย เซลล์ไข่ (Egg Cell) ซึงมีขนาดใหญ่อยู่ตรงกลาง และ 2 Synergid Cell อยู่ 2 ข้างของ ่ เซลล์ไข่ 2 นิวเคลียสทีอยู่ตรงกลาง คือ Polar Nuclei ซึ่งต่อมาจะรวมกันเป็ น 2 n ่ โครโมโซม และเปลียนเป็ น Endosperm Mother Cell ดังนันเมือสินสุดกระบวนการสร้าง ่ ้ ่ ้ 452 CU 474
41.
เซลล์สบพันธุเพศเมีย ภายใน Embryo
Sac จึงมีเซลล์ทงหมด 7 เซลล์ ทีเปลียนแปลงมาก ื ์ ั้ ่ ่ จาก 8 นิวเคลียส โดยมีเซลล์ไข่ (Egg Cell) เป็ นเซลล์สบพันธุเพศเมีย (Female Gamete) ื ์ รูปที่ 10.3.5 การสร้างเซลล์สบพันธุเพศเมียของพืชดอก ื ์ การถ่ายละอองเรณู (Pollination) การถ่ายละอองเรณู คือ การทีละอองเรณูตกลงบนยอดเกสรตัวเมีย โดยมีขนหรือ ่ สารเหนียวทียอดเกสรตัวเมีย เป็ นตัวช่วยยึดจับเอาไว้ ่ เมือละอองเรณูแก่เต็มที่ ผนังของ Pollen Sac จะฉีกขาดพร้อมกับอับเรณูแตกออก ่ ทําให้ละอองเรณูปลิวออกมา การถ่ายละอองเรณูจาเป็ นต้องอาศัยสือ (Pollinating Agent) ํ ่ พาไป ซึงมีหลายชนิด ได้แก่ ่ ลม พืชทีละอองเรณูตองอาศัยลมพาไป มักมีดอกขนาดเล็ก กลีบดอกไม่สวย ไม่มส ี ่ ้ ี ไม่มต่อมนํ้าหวานทีจะดึงดูดแมลง ดอกพวกนี้ละอองเรณูแห้งและเบาบาง ชนิดมีปีกช่วยให้ ี ่ ปลิวไปได้ไกล ขณะเดียวกัน ยอดเกสรตัวเมียของพืชทีตองอาศัยลมในการถ่ายละอองเรณู ่ ้ มักจะมีโครงสร้างช่วยยึดเกาะ เช่น มีขนมาก มีกงแตกเป็ นแขนงเล็ก ๆ มีสารเหนียว เป็ น ิ่ ต้น พืชพวกนี้ ได้แก่ หญ้า ข้าว CU 474 453
42.
แมลง แมลงมากชนิดอาศัยนํ้ าหวานจากดอกไม้เป็
นแหล่งอาหาร การเข้ามาดูด นํ้าหวานทําให้ละอองเรณูตดตัวไปตกบนยอดเกสรตัวในดอกเดียวกันหรืออื่น ๆ ได้ แมลง ิ ทีเป็ นสือมากทีสด คือ ผึงและผีเสือ จึงได้ชอว่าเป็ นแมลงนักผสมพันธุพช ่ ่ ุ่ ้ ้ ่ื ์ ื นอกจากนี้ ยังมีน้ําเป็ นสือในการถ่ายละอองเรณูทสาคัญของพืชนํ้า รวมถึงสัตว์อ่น ๆ ่ ่ี ํ ื เช่น นก หอยทาก ก็มสวนช่วยในการถ่ายละอองเรณู ี่ รูปที่ 10.3.6 การถ่ายละอองเรณูโดยมีแมลงเป็ นสือ ่ การถ่ายละอองเรณูของพืช เกิ ดได้ 2 แบบ คือ 1. Self Pollination คือ การถ่ายละอองเรณูในดอกเดียวกัน หรือต่างดอกในต้น เดียวกัน หรือ ต่างดอกต่างต้นทีเป็ นพันธุบริสทธิ ์ ในพืชทีเป็ นสายพันธุเดียวกัน ่ ์ ุ ่ ์ 2. Cross Pollination คือ การถ่ายละอองเรณูในพืชต่างต้นทีมคุณสมบัตทาง ่ ี ิ กรรมพันธุต่างกัน โดยเป็ น Species เดียวกัน หรือต่าง Species ต่าง Genus กันก็ได้ ์ การปฏิ สนธิ (Fertilization) เมือละอองเรณูตกลงบนยอดเกสรเพศเมีย จะได้รบความชืนและอาหารพวกนํ้าตาล ่ ั ้ จากยอดเกสรเพศเมีย งอกหลอดเรณู (Pollen Tube) อัตราความเร็วในการงอกแตกต่าง กันในพืชแต่ละชนิด ซึงอาจใช้เวลาเป็ นวัน สัปดาห์ หรือเดือน หลังการถ่ายละอองเรณูกได้ ่ ็ หลอดเรณูจะแทงลงไปในเนื้อเยือของยอดเกสรเพศเมีย ผ่านไปตามก้านเกสรเพศเมีย เพือ ่ ่ นํา Generative Nucleus ไปทีโอวุล เมือหลอดเรณูมาถึงโอวุล จะเข้าสู่ Embryo Sac ทาง ่ ่ ไมโครไพล์ Tube Nucleus จะสลายไปพร้อมกับปลายหลอดเรณูแตกออก ซึงก่อนหน้านี้ ่ Generative Nucleus จะแบ่งตัวแบบไมโตซิส ได้เป็ น 2 Nuclei ทีเรียกว่า Sperm Nucleus ่ เมื่อปลายหลอดเรณูแตกออก Sperm Nucleus ทัง 2 จะเข้าสู่ Embryo Sac Sperm ้ Nucleus หนึ่งจะผสมกับไข่ได้เป็ นไซโกต (Zygote) (2n) Sperm Nucleus อีกอันหนึ่งจะ ผสมกับ Polar Nuclei ได้เป็ น Primary Endosperm Cell (3n) ไซโกตจะเจริญต่อไปเป็ น Embryo ในขณะที่ Primary Endosperm Cell เจริญต่อไปเป็ น Endosperm Tissue ทํา 454 CU 474
43.
หน้าทีสะสมอาหารให้ Embryo ส่วน
Antipodal และ Synergid Cell จะสลายตัวไป จะเห็น ่ ว่ า มี ก ารเกิ ด การผสม 2 ครัง เรี ย กการผสมพัน ธุ์ น้ี ว่ า การปฏิ ส นธิ ซ้ อ น (Double ้ Fertilization) ซึงจะเกิดขึนกับพืชดอกเท่านัน ่ ้ ้ รูปที่ 10.3.7 การปฏิสนธิในพืชดอก (จาก โครงการตําราวิทยาศาสตร์และคณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 1) การเจริญเติบโตของพืชหลังการปฏิสนธิ หลังจากเกิดการปฏิสนธิแล้ว โอวุล หรือ ถุงเอมบริโอ (Embryo Sac) จะเจริญไป เป็ นเมล็ด (Seed) โดยรังไข่เจริญไปเป็ นผล (Fruit) ซึงมีเนื้อเยื่อรังไข่ทห่อหุมโอวุลเป็ นเนื้อ ่ ่ี ้ ของผลหุมเมล็ด ไซโกตจะแบ่งเซลล์เติบโตและเปลียนแปลงไปเป็ นกลุ่มเนื้อเยื่อ สร้างรูป ้ ่ เป็ นตัวอ่อน (Embryo) โดยมี Primary Endosperm เปลียนแปลงเป็ นเนื้อเยื่อเอนโด่ สเปอร์ม สะสมอาหารในเมล็ดเพือให้ตนอ่อนใช้ในขณะงอกเป็ นต้นกล้า ่ ้ โครงสร้างของผล (Structure of Fruit) รังไข่ทเี่ จริญมาเป็ นผล จะมีผนังรังไข่ เจริญขึนมาเป็ นผนังของผล หรือ เปลือกของ ้ ผล (Pericarp) ซึ่งอาจมีความหนามากน้อยต่างกันตามชนิดของผล บางชนิดอาจเป็ น เนื้อเยือทีรบประทานได้ เปลือกของผลแบ่งเป็ น 3 ชัน ได้แก่ ่ ่ั ้ 1. เปลือกชันนอก (Exocarp) ส่วนใหญ่เป็ นเนื้อเยื่อเอพิเดอร์มสเพียงชันเดียว ้ ิ ้ เป็ นชันบาง ๆ แต่ผลบางชนิดก็มหลายชัน ผลบางชนิดอาจมีหนาม ขน และบางชนิดมี ้ ี ้ ต่อมนํ้ามัน มีปากใบ 2. เปลือกชันกลาง (Mesocarp) เป็ นเนื้อเยื่อบาง ๆ ประกอบด้วย Vascular ้ Bundle บางชนิดเป็ นเนื้อเยืออ่อนนุ่มรับประทานได้ แต่บางชนิดแข็งรับประทานไม่ได้ ่ CU 474 455
44.
3. เปลือกชันใน (Endocarp)
เป็ นเนื้อเยื่อที่ประกอบด้วย เซลล์ชนเดียว หรือ ้ ั้ หลายชัน บางชนิดเป็ นเนื้อนุ่มรับประทานได้ ้ Pericarp ของผลบางชนิด อาจแยกเป็ นชันต่าง ๆ เห็นได้ชด เช่น มะม่วง มะพร้าว ้ ั พุทรา เป็ นต้น แต่ผลบางชนิดชันทัง 3 ของ Pericarp เชือมติดกันจนแยกไม่ออก เช่น ้ ้ ่ ข้าวโพด พริก Pericarp ของผลบางชนิดแห้งและแข็ง เช่น ถัวลิสง ถัวเหลือง บางชนิดแข็ง ่ ่ และเหนียว เช่น เปลือกและกะลามะพร้าว ชนิ ดของผล ผลแบ่งได้เป็ น 3 กลุมใหญ่ ๆ ได้แก่ ่ 1. ผลเดียว (Simple Fruit) คือ ผลทีเปลียนแปลงมาจากรังไข่เดียวของดอกเดียว ่ ่ ่ ่ หรือดอกช่อ ใน 1 รังไข่น้ีอาจประกอบด้วย คาร์เพล (Carpel) เดียวหรือหลายคาร์เพลที่ เชื่อมติดกัน เช่น มะเขือ องุ่น ละมุด กล้วย ฝรัง ฟก แตง เป็ นต้น ผลเดี่ยวจําแนกตาม ่ ั ลักษณะของ Pericarp ได้เป็ น 2 ชนิด คือ ผลสด (Fleshy Fruit) คือ ผลทีม ี Pericarp ่ ส่วนใหญ่หรือทังหมดเป็ นเนื้ออ่อนนุ่ม และผลแห้ง (Dry Fruit) คือ ผลชนิดทีมเี ปลือกแห้ง ้ ่ เมือผลแก่ซงเปลือกอาจจะแตกได้เองหรือไม่แตก ่ ่ึ 2. ผลกลุ่ม (Aggregate Fruit) คือ ผลทีเปลียนแปลงมาจากกลุ่มของรังไข่ทอยู่ใน ่ ่ ่ี ดอกเดียวทีอยู่บนฐานรองดอกเดียวกัน รังไข่แต่ละอันเป็ นผลย่อยหนึ่งผลอยู่อดกันแน่นจน ่ ่ ั เห็นเป็ นผลเดียวขนาดใหญ่ เช่น น้อยหน่ า ผลบางอย่างผลย่อยแยกกันเป็ นผลเล็ก ๆ เช่น ่ กระดังงา การะเวก บางชนิดผลคือส่วนของฐานรองดอก เช่น สตรอเบอรี่ ฐานรองดอก เติบโตขึนมากลายเป็ นเนื้อรับประทานได้ โดยมีผลทีแท้จริงเป็ นผลย่อย ๆ ติดอยู่ใกล้กบผิว ้ ่ ั นอก 3. ผลรวม (Multiple Fruit) คือ ผลทีเปลียนแปลงมาจากกลุ่มรังไข่ของดอกทังช่อ ่ ่ ้ เชื่อมรวมกันแน่ น รังไข่ท่ีเจริญเป็ นผลย่อยจะเชื่อมรวมกันแน่ นจนเป็ นผลเดีย ว ได้แ ก่ สับปะรด ขนุน สาเก ยอ หม่อน มะเดือ ่ 456 CU 474
45.
ก ข ค รูปที่ 10.3.8 ชนิดของผล
ผลเดียว (ก) ผลกลุม (ข) และผลรวม (ค) ่ (จาก โครงการตําราวิทยาศาสตร์และคณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 1) เมล็ด (Seed) เมล็ด คือ โอวุลทีแก่แล้วเจริญเติบโตขึนหลังจากการปฏิสนธิ มีสวนประกอบ ดังนี้ ่ ้ ่ 1. เปลือกเมล็ด (Seed Coat) เป็ นส่วนทีอยู่ชนนอก เกิดจากผนังโอวุล (Integument) ่ ั้ ทําหน้าทีป้องกันอันตรายและการสูญเสียความชืนจากเนื้อเยื่อของเมล็ด ส่วนมากมี 2 ชัน ่ ้ ้ คือ ชันนอก (Testa) เปลียนแปลงมาจาก Outer Integument มักหนา เหนียว และชันใน ้ ่ ้ (Tegmen) เปลียนแปลงมาจาก Inner Integument เป็ นเยื่อบาง ๆ บนเปลือกเมล็ดมีรอย ่ แผลเป็ น (Hilum) ทีมรไมโครไพล์ ซึงเป็ นทางผ่านของ Pollen Tube มาก่อน เปลือกเมล็ด ่ ีู ่ อาจมีการเปลียนแปลงไปในพืชบางอย่าง เช่น เปลือกเมล็ดชันนอกของทุเรียน ลินจี่ ลําไย ่ ้ ้ เงาะ เปลียนเป็ นเนื้อนุ่ มหุมเมล็ด เป็ นส่วนทีรบประทานได้ ในพวกแคแสด กระเทียมเถา ่ ้ ่ั เปลียนแปลงเปลือกเมล็ดเป็ นแผ่นบาง ๆ ติดกับเมล็ด ทําให้ปลิวไปได้ไกล ช่วยในการแพร่ ่ พันธุ์ เป็ นต้น 2. เอมบริโอ (Embryo) เป็ นส่วนทีเจริญมาจากไซโกต ประกอบด้วย 3 ส่วน ่ 1) ใบเลียง (Cotyledon) มีหน้าทีสะสมอาหารไว้เลียงต้นอ่อน โดยทําการย่อย ้ ่ ้ และดูดซึมอาหารที่อยู่รอบ ๆ คือ เอนโดสเปอร์มมาเก็บไว้ พืช บางชนิดใบเลี้ย งสะสม อาหารไว้ม าก ทํ า ให้ใ บเลี้ย งมีล ัก ษณะหนา ในเมล็ด จึง ไม่ ม ีเ อนโดสเปอร์ม เหลือ อยู่ พืชบางชนิดใบเลียงสะสมอาหารมากใบจึงบาง เมล็ดจะมีเอนโดสเปอร์มมาก นอกจากนี้ใบ ้ CU 474 457
46.
เลียงยังป้องกันยอดอ่อนและช่วยในการสังเคราะห์ดวยแสง เพื่อเลียงต้นกล้าในระยะทียง ้ ้ ้ ่ ั ไม่มใบแท้
เพราะใบเลียงมีคลอโรฟิล ี ้ 2) ลําต้นอ่อน (Caulicle) เป็ นแกนกลางมี 2 บริเวณ คือ บริเวณที่อยู่เหนือ ใบเลียงขึนไป เรียกว่า เอพิคอทิล (Epicotyl) ซึงบริเวณปลายสุดเป็ นส่วนของยอดแรกเกิด ้ ้ ่ (Plumule) ในพืชใบเลียงเดียวจะมีเยื่อบาง ๆ หุม เรียกว่า Coleoptile ส่วนบริเวณทีอยู่ใต้ ้ ่ ้ ่ ใบเลียงลงมา เรียกว่า ไฮโปคอทิล (Hypocotyl) ซึงปลายสุดเป็ นรากแรกเกิด (Radicle) ซึง ้ ่ ่ จะเจริญไปเป็ นรากแก้ว (Primary Root) ต่อไป ในพืชใบเลียงเดียวจะมีเยื่อบาง ๆ หุมราก ้ ่ ้ แรกเกิด เรียกว่า Coleorhiza 3. เอนโดสเปอร์ม (Endosperm) เป็ นเนื้อเยื่อของเมล็ดทีเจริญมาจาก Primary ่ Endosperm Cell (3n) เพือสะสมอาหารไว้ให้ตนอ่อนใช้ในระยะแรกของการงอก อาหารที่ ่ ้ สะสมมีทงคาร์โบไฮเดรต ไขมัน โปรตีน และสารอื่น ๆ มีทงชนิดที่เป็ นของเหลว (Liquid ั้ ั้ Endosperm) เช่น นํ้ ามะพร้าว นํ้ าลูกตาล นํ้ าลูกหมาก และชนิดทีเป็ นของแข็ง (Fleshy ่ Endosperm) เช่น เนื้อมะพร้าว เนื้อลูกตาล เนื้อลูกหมาก เป็ นต้น ก. ข. รูปที่ 10.3.9 ส่วนประกอบของเมล็ด : เมล็ดพืชใบเลียงคู่ (ก) เมล็ดพืชใบเลียงเดียว (ข) ้ ้ ่ การเจริ ญระยะเอมบริ โอของพืชดอก การเติบโตของเอมบริโอเกิดอยู่ภายใน โดยเริมจากไซโกตแบ่งไมโตซิสครังแรกได้ ่ ้ 2 เซลล์ เซลล์ทอยูตดไมโครไพล์เรียกว่า เบซัล (Basal Cell) และเซลล์ทอยู่ห่างไมโครไพล์ คือ ่ี ่ ิ ่ี เทอร์มนล (Terminal Cell) เซลล์เบซัลจะแบ่งไมโตซิสอีกหลายครังในแนวเดียว เซลล์เรียง ิ ั ้ ตัวเป็ นแถวเดียวยาว โดยเซลล์ล่างสุดขยายใหญ่กว่าเซลล์อ่น ๆ ทีแบ่งได้น้ีเป็ น ซัสเปน ื ่ เซอร์ (Suspensor) สําหรับยึดเอมบริโอให้ตดกับ Embryo Sac ส่วนเซลล์เทอร์มนลจะ ิ ิ ั แบ่ ง ตัว เป็ น 2 แนวทาง คือ แนวตัง และแนวนอน ทํา ให้ไ ด้ก ลุ่ ม เซลล์เ ป็ น รูป ทรงกลม ้ 458 CU 474
47.
(Globular) อยู่ท่ปลายสุดของซัสเปนเซอร์ กลุ่มเซลล์น้ีจะพัฒนาไปเป็ นเอมบริโอ ี (Globular
Embryo) โดยเซลล์จะเติบโตและจัดกลุ่มเป็ นเนื้อต่าง ๆ สร้างรูปของต้นอ่อน ที่ ประกอบด้วย ใบเลียงและต้นอ่อน ้ ก. ข. รูปที่ 10.3.10 การเจริญของต้นอ่อนในเมล็ดพืชใบเลียงคู่ (ก) และพืชใบเลียงเดียว (ข) ้ ้ ่ การงอกของเมล็ด (Seed Germination) การงอกของเมล็ด คือ การกลับคืนสู่สภาพการเจริญเติบโตของเมล็ดโดยเอมบริโอพ้น จากระยะการพักตัว (Dormancy) เริมมีกจกรรมของชีวตเจริญงอกออกมาจากเมล็ดและ ่ ิ ิ เปลียนแปลงเป็ นต้นใหม่ ่ ั การงอกของเมล็ดต้องการปจจัยทีจาเป็ น ได้แก่ ่ํ 1. นํ้า เมล็ดจะงอกได้ตองอยู่ในสภาพทีมความชืนทีพอเหมาะ นํ้าช่วยทําให้เปลือก ้ ่ ี ้ ่ เมล็ดอ่อนนุ่ ม ทําให้ตนอ่อนสามารถดันทะลุเปลือกออกมาได้ ช่วยทําให้ออกซิเจนผ่านเข้า ้ สู่เมล็ดได้ง่าย นํ้ าที่เข้าสู่เมล็ดช่วยละลายสารในโปรโตพลาสซึมเจือจางลงจนทํางานได้ และช่วยในการนํ าสารอาหารทีสะสมจากเอนโดสเปอร์มไปเลียงส่วนต่าง ๆ ของเอมบริโอ ่ ้ ได้ 2. ออกซิเจน การงอกของเมล็ดต้องใช้พลังงานเพื่อการแบ่งเซลล์ การเพิมขนาด ่ ของเซลล์จงจําเป็ นต้องใช้ออกซิเจนในการเผาผลาญอาหารที่สะสมอยู่ในเมล็ด พลังงาน ึ CU 474 459
48.
บางส่วนจะเปลียนเป็ นความร้อน ดังนันขณะเมล็ดงอกจึงเกิดความร้อนด้วย
เมล็ดจะงอก ่ ้ ได้ดทปริมาณออกซิเจน 20 เปอร์เซ็นต์หรือมากกว่า สําหรับพืชนํ้าทีมปริมาณออกซิเจนตํ่า ี ่ี ่ ี แต่สามารถงอกได้ดี เพราะเมล็ดหายใจแบบไม่ใช้ออกซิเจน (Anaerobic Respiration) 3. อุณหภูม ิ พืชแต่ละชนิดต้องการอุณหภูมสาหรับการงอกของเมล็ดทีแตกต่างกัน ิํ ่ พืช เขตร้อ นต้อ งการอุ ณ หภู ม ิสูง กว่า พืชเขตหนาว แต่ อุ ณ หภู ม ิท่ีพ อเหมาะ (Optimum Temperature) ของเมล็ดพืชทัวไปอยูระหว่าง 20-30 องศาเซลเซียส ่ ่ ั นอกจากนี้พชบางชนิดอาจต้องการปจจัยอย่างอื่นสําหรับการงอกของเมล็ด เช่น ื พืชพวกหญ้า และยาสูบ เมล็ดจะงอกได้ดถ้าได้รบแสงสว่าง โดยทัวไปแสงสว่างไม่เป็ น ี ั ่ ั ปจจัยสําหรับการงอกของเมล็ดพืชทัวไป เนื่องจากยังไม่มกระบวนการสังเคราะห์ดวยแสง ่ ี ้ ั ยิงกว่านันแสงสว่างอาจเป็ นปจจัยยับยังการงอกของเมล็ด เช่น หอมใหญ่ เป็ นต้น ่ ้ ้ ลักษณะการงอกของเมล็ด การงอกของเมล็ดจําแนกได้ 2 ลักษณะ ได้แก่ 1. Epigeal Germination เป็ นการงอกโดยรากแรกเกิด (Radicle) จะงอกออกมา ก่อน กลายเป็ นรากแก้ว จากนันส่วนของลําต้นใต้ใบเลี้ยง (Hypocotyl) จะเจริญและ ้ ยืดตัวอย่างรวดเร็ว ดึงใบเลียงและลําต้นเหนือใบเลียง (Epicotyl) ออกจากเปลือกเมล็ดชู ้ ้ ขึนเหนือดิน เมื่อใบเลี้ยงกางออก ลําต้นเหนือใบเลี้ยงจะยืดตัวตรงขึนชูส่วนยอดแรกเกิด ้ ้ (Plumule) เมือเกิดใบแท้ทสงเคราะห์ดวยแสงได้ ซึงเป็ นเวลาเดียวกับใบเลียงถูกใช้อาหาร ่ ่ี ั ้ ่ ้ หมดไปจึงเหียวและหลุดไป พืชทีมการงอกในลักษณะนี้ ได้แก่ พืชใบเลี้ยงคู่ และพวกถัว ่ ่ ี ่ เมล็ดยาว 2. Hypogeal Germination เป็ นการงอกทีสวนของต้นอ่อนใต้ใบเลียง (Hypocotyl) ่่ ้ เจริญเติบโตช้ากว่าลําต้นเหนือใบเลียง (Epicotyl) ดังนัน ลําต้นเหนือใบเลียงกับยอดแรก ้ ้ ้ เกิดเท่านันทีงอกขึนมาเหนือพืนดิน โดยมีส่วนลําต้นใต้ใบเลียงจมอยู่ใต้ดน ส่วนใหญ่เป็ น ้ ่ ้ ้ ้ ิ การงอกของพืช ใบเลี้ย งเดี่ย วพวกข้า ว ข้า วโพด หญ้ า ส่ ว นพวกใบเลี้ย งคู่ เป็ น พวก ถัวเมล็ดกลม (Pea) เช่น ถัวลันเตา และพืชพวกส้ม ่ ่ 460 CU 474
49.
ก. ข. รูปที่ 10.3.11 การงอกของเมล็ดชนิดต่าง
ๆ พืชใบเลียงคู่ (ก) และพืชใบเลียงเดียว (ข) ้ ้ ่ (จาก โครงการตําราวิทยาศาสตร์และคณิตศาสตร์ มูลนิธิ สอวน. 2551, ชีววิทยา 1) ระยะพักตัวของเมล็ด (Seed Dormancy) ระยะพักตัว คือ ช่วงเวลาที่พชหยุดการเจริญชัวคราว ไม่มกจกรรมการดํารงชีพ ื ่ ี ิ ทังนี้เพื่อหลีกเลี่ยงสภาพแวดล้อมที่ไม่เหมาะสม หรือเกิดจากสภาวะภายในของพืชที่ไม่ ้ พร้อมจะทํางาน เมล็ดพืชบางชนิดแม้จะแก่แล้วแต่ย งต้องรอเวลาที่จะงอก ซึ่งเกิดจาก ั สภาพภายในของเมล็ด การพักตัวของเมล็ดมีหลายสาเหตุ ได้แก่ 1. เปลือกเมล็ด เมล็ดพืชบางชนิดมีเปลือกแข็งและหนามาก เป็ นอุปสรรคต่อการ ขยายตัวของ Embryo ทัง ๆ ทีน้ําและออกซิเจนสามารถซึมผ่านได้ดี ทําให้เมล็ดพักตัวอยู่ ้ ่ ได้นานหลายสิบปี เช่น เมล็ดผักกาด และผักขมบางชนิด เมล็ดบางอย่างเปลือกเมล็ดไม่ ยอมให้น้ําซึมผ่าน เช่น เมล็ดพืชตระกูลถัว และผักบุงฝรัง่ โดยเฉพาะอย่างยิงถ้าเก็บไว้ใน ่ ้ ่ ทีแห้งนํ้ าจะซึมผ่านได้ชามาก แต่ถ้าอยู่ในธรรมชาติในดินมีจุลนทรียช่วยย่อยทําให้น้ํ าซึม ่ ้ ิ ์ ผ่านเปลือกเมล็ดได้ดขน ระยะพักตัวก็จะน้อยลง ทําให้งอกได้เร็วขึน ี ้ึ ้ เมล็ดบางชนิดมีเยื่อหุม Embryo ทําให้ Embryo ไม่ได้รบออกซิเจน การงอกจึงช้า ้ ั ลง เช่น เมล็ดกระจับ และหญ้าชนิดต่าง ๆ 2. การเจริญของเอมบริโอในเมล็ด ปกติเมล็ดเมื่อแก่จะแห้งและหลุดออกจากต้น เอมบริโอจะเจริญเต็มที่พร้อมที่จะงอก แต่เมล็ดบางอย่างเอมบริโอยังต้องใช้เวลาในการ เจริญอีกระยะหนึ่งภายหลังเมล็ดแก่ ซึ่งอาจนานเป็ นสัปดาห์ห รือเป็ นเดือน เช่น เมล็ด แป๊ะก๊วย (Ginkgo biloba) และกล้วยไม้บางชนิด CU 474 461
50.
3. การพักตัวของเอมบริโอ (Dormant
Embryo) เมล็ดพืชบางชนิดมีการพักตัวใน ระยะเอมบริโอ โดยต้องการระยะเวลาในการเปลี่ยนแปลงทางสรีรวิทยาของเนื้อเยื่อให้ สมบูรณ์ ดังนันแม้ว่าเอมบริโอเจริญเต็มที่แล้วอยู่ในสภาพทีพร้อมจะงอก คือ มีความชื้น ้ ่ และออกซิเจน ปราศจากเปลือกเมล็ดแล้วก็ตาม เมล็ดก็จะยังไม่งอก เช่น แอปเปิ้ ล ท้อ สน สองใบ สนสามใบ ต้องการอุณหภูมต่ํา ความชืนสูง เป็ นระยะยาวนานในการเปลียนแปลง ิ ้ ่ สรีรวิทยาของเนื้อเยือ เอมบริโอจึงมักพักตัวตลอดช่วงเวลาฤดูหนาว ่ 4. สารเคมีทยบยังการงอก (Germination Inhibitor) เมล็ดของพืชบางชนิดมีสาร ่ี ั ้ ยับยังการงอกของเมล็ด พวกกรดแอบไซซิก (ABA) เคลือบอยู่ เช่น มะเขือเทศ มะละกอ มี ้ เมือกทีหุมเมล็ดเป็ นสารยับยังการงอกของเมล็ด ดังนัน ก่อนนําเมล็ดไปเพาะควรล้างเมือก ่ ้ ้ ้ หรือนําไปตากแดดให้แห้ง จะทําให้เมล็ดงอกเร็วขึน ้ นอกจากนี้ยงมีสาเหตุอ่น ๆ ที่ทําให้เกิดการพักตัว แต่ไม่ใช่สาเหตุท่เกิดโดยตรง ั ื ี เช่น แสง อาจเป็ นตัวยับยังการงอกของเมล็ดบางชนิด ความชืน พืชบางชนิดงอกได้ดทม ี ้ ้ ี ่ี ความชืนสูง แต่พชบางชนิดเมล็ดทีเก็บมาใหม่ ๆ ความชืนสูงจะไม่งอกต้องรอให้เมล็ดแห้ง ้ ื ่ ้ จึงจะงอก และอุณหภูม ิ พบว่า พืชเขตหนาว จะพักตัวในช่วงฤดูหนาว ส่วนพืชเขตร้อนจะ พักตัวในฤดูรอน ้ ความสามารถในการงอกของเมล็ด (Seed Viability) ความสามารถในการงอกของเมล็ด (Seed Viability) คือ ความสามารถของเมล็ด ในการมีชวตที่จะงอกเป็ นต้นอ่อนหรือกล้า (Seedling) ได้ เมล็ดของพืชแต่ละชนิดมี ีิ ความสามารถในการงอกในระยะที่แตกต่างกัน บางชนิดถ้าเก็บไว้นานเกินไปจะไม่งอก ความสามารถในการงอกของเมล็ดขึนอยูกบปจจัยต่าง ๆ ได้แก่ ้ ่ ั ั 1. การเจริญเติบโตเต็มทีของเมล็ด (Maturity of Seed) เมล็ดทีแก่จดจะมี ่ ่ ั ความสามารถในการงอกสูงกว่าเมล็ดทียงไม่แก่ และต้นอ่อนทีงอกออกมาจะแข็งแรงกว่า ่ ั ่ 2. ความแข็งแรงของต้นพันธุ์ ถ้าต้นพันธุมความแข็งแรง ปราศจากโรค เมล็ดสะสม ์ ี อาหารมาก เอมบริโอมีขนาดใหญ่ ต้นกล้าทีงอกออกมาก็จะแข็งแรง ่ 3. การเก็บรักษาเมล็ดพืช วิธการเก็บรักษาเมล็ดพืช ต้องเก็บในสภาพทีมอุณหภูม ิ ี ่ ี ตํ่า ความชื้นน้อย จะทําให้รกษาความสามารถในการงอกของเมล็ดไว้ได้นาน ปกตินิยม ั เก็บทีอุณหภูมประมาณ 5 องศาเซลเซียส ่ ิ 462 CU 474
51.
4. อายุของเมล็ด (Longevity)
เมล็ดของพืชแต่ละชนิดมีอายุแตกต่างกันมาก เช่น เมล็ด บัว มีอ ายุ ถึง 400 ปี เมล็ด พืช ตระกู ล ถัว บางชนิ ด มีอ ายุ ถึง 75 ปี ในขณะที่เ มล็ด ่ กล้วยไม้มอายุเพียง 2-3 สัปดาห์เท่านัน ี ้ ความแข็งแรงของเมล็ด ความแข็งแรงของเมล็ด หมายถึง เมล็ดที่มความสามารถในการงอกได้รวดเร็ว ี สมํ่าเสมอ และต้นกล้าสามารถตังตัวเร็วเมื่อนํ าไปปลูก การตรวจสอบคุณภาพของเมล็ด ้ พันธุพช เพือพิจารณาความแข็งแรงของเมล็ดพันธุ์ ทําได้หลายวิธี ได้แก่ ์ ื ่ 1. การเร่งอายุเมล็ด เป็ นวิธการตรวจสอบความแข็งแรงของเมล็ด เพื่อทํานายว่า ี เมื่อ เมล็ด เก็บ ไว้เ ป็ น เวลานาน ค่ า การงอกของเมล็ด จะยัง คงสูง อยู่ ห รือ ไม่ ทํา โดยนํ า ตัวอย่างเมล็ดจากแหล่งทีตองการตรวจสอบมาให้ความร้อนในตูอบทีอุณหภูม ิ 40-45 องศา ่ ้ ้ ่ เซลเซียส ความชืนสัมพัทธ์ ร้อยละ 100 เป็ นเวลา 2-3 วัน แล้วนําเมล็ดไปทําการเพาะหา ้ ค่าร้อยละของการงอก ถ้าพบว่ามีอตราการงอกสูง แสดงว่าเมล็ดพันธุจากแหล่งนันมีความ ั ์ ้ แข็ง แรง ทํ า นายได้ว่ า เมล็ด พัน ธุ์ ส ามารถเก็บ ไว้ไ ด้โ ดยไม่ ต้อ งควบคุ ม อุ ณ หภู ม ิ และ ความชืนสัมพัทธ์เป็ นเวลานาน 12-18 เดือน ้ การเร่งอายุ เมล็ดใช้หลักการทีทาให้องค์ประกอบทางเคมีในเมล็ดเสือมสภาพอย่าง ่ ํ ่ รวดเร็ว เช่นเดียวกับเมล็ดทีเก็บในสภาพปกติเป็ นเวลานาน การเร่งอายุเมล็ดจึงเป็ นการ ่ ทําให้เมล็ดเสือมสภาพ เสมือนว่าเมล็ดนันถูกเก็บไว้เป็ นเวลานานนันเอง ่ ้ ่ 2. การวัดดัชนีการงอกของเมล็ด เป็ นวิธทใช้หลักการความแข็งแรงของเมล็ด คือ ี ่ี เมล็ดทีมความแข็งแรงสูงย่อมจะงอกได้เร็วกว่าเมล็ดทีมความแข็งแรงตํ่า โดยนํ าตัวอย่าง ่ ี ่ ี เมล็ดพันธุ์แหล่งต่าง ๆ มาเพาะแล้วนับจํานวนเมล็ดทีงอกในแต่ละวันนํ ามาคํานวณหาค่า ่ ดัชนีการงอก ดังนี้ จํานวนเมล็ดทีงอกในแต่ละวัน ่ ดัชนีการงอก = ผลบวกของ จํานวนวันหลังเพาะ CU 474 463
52.
ตัวอย่างเช่น เมล็ดถัวเหลือง ตัวอย่างที่
1 จํานวน 100 เมล็ด เริมทําการเพาะในวันที่ 1 ่ ่ บันทึกข้อมูลการงอกได้ดงนี้ ั วันทีเพาะ ่ เมล็ด 1 2 3 4 5 6 7 จํานวนเมล็ดที่ งอก 42 33 15 - = 42 + 33 + 15 4 5 3 = 14 + 8.25 + 3 = 25.25 นําค่าดัชนีการงอกของเมล็ดพันธุ์จากแหล่ง ต่าง ๆ มาเปรี ยบเทียบกัน เมล็ดพันธุ์ที่มีค่าดัชนี การงอกสูง แสดงว่าเมล็ดพันธุ์จากแหล่งนั้น แข็งแรงกว่าแหล่งอื่น ดัชนีการงอก 10.4 การตอบสนองของพืช (Plant Response) การตอบสนองของพืชต่อสิ่งแวดล้อม พืช ไม่ส ามารถเคลื่อ นที่ไ ด้อ ย่ า งสัต ว์เ นื่ อ งจากพืช มีร ากที่ห ยัง ลึก ลงดิน เพื่อ ยึด ่ ั เหนี่ยวลําต้น แต่ก็สามารถเคลื่อนไหว (movement) เพื่อตอบสนองต่อปจจัยสิงแวดล้อม ่ ทีมากระตุนได้ ่ ้ การเคลื่อนไหวของพืชที่ตอบสนองต่อปัจจัยภายนอก ั การตอบสนองต่ อ ป จ จัย ภายนอกของพืช ช่ วยให้พ ืชมีก ารเจริญ เติบ โตได้อ ย่ า ง เหมาะสมกับสภาพแวดล้อมของพืช การเคลือนไหวของพืชแบ่งได้ 2 ประเภท ่ ั 1. การตอบสนองอย่ า งมี ทิ ศ ทางต่ อ ป จ จัย สิ่ ง แวดล้ อ มที่ ม ากระตุ้ น (tropic movement หรือ tropism) เป็ นการเคลื่อนไหวของพืชทีเกิดจากการเจริญเติบโตอย่างมี ่ ั ทิศทางที่ตอบสนองต่อปจจัยภายนอก ซึ่งเป็ นตัวกําหนดทิศทางการเคลื่อนไหวของพืช เช่น การตอบสนองต่อแรงโน้มถ่วงของโลก (gravitropism) การตอบสนองต่อทิศทางของ ั แสง (phototropism) ถ้าตอบสนองโดยการเคลื่อนไหวเข้าหาปจจัยกระตุนเรียกว่า positive ้ ั tropism ในกรณีทพชเคลือนไหวหนีออกจากปจจัยกระตุนเรียกว่า negative tropism ่ี ื ่ ้ 464 CU 474
53.
ั 2. การตอบสนองอย่า งไม่ม
ีทิศ ทางที่แน่ น อนต่ อ ป จ จัย สิง แวดล้อ มที่ม ากระตุ้น ่ ั (nastic movement) เป็ นการเคลื่อนไหวของพืชทีไม่ถูกกําหนดด้วยทิศทางของปจจัย ่ ั ภายนอกที่มากระตุ้น เช่น การหุบบานของดอก ซึ่งมีปจจัยที่สําคัญ ได้แก่ แสง (Light) และอุณหภูม ิ (Temperature) กลไกการตอบสนองของพืชอาจเกิดจากการเจริญเติบโตที่ แตกต่างกันของส่วนต่าง ๆ หรือมีการเปลียนแปลงความเต่งของเซลล์พเิ ศษ ่ กลไกทีทาให้เกิดการเคลือนไหวของพืชเชือว่าเป็ นผลจาก ่ ํ ่ ่ 1. การเจริญเติบโตทีไม่เท่าเทียมกัน (different growth) ของพืชซึงส่วนใหญ่จะไม่ ่ ่ ย้อนกลับมาได้อก ี 2. การเคลื่อนของนํ้ าเข้าไปในเซลล์พเศษที่ทําหน้ าที่เกี่ยวกับการเคลื่อนไหว คือ ิ motor cells ทีรวมกันเป็ นเนื้อเยื่อ พัลวินัส (pulvinus) กลุ่มเซลล์พวกนี้เป็ นพวกเซลล์ ่ parenchyma ทีมหนังบางและมีขนาดใหญ่ นํ้ าจะผ่านเข้าออกแต่ละเซลล์ไม่เท่ากัน เมื่อ ่ ี เซลล์มน้ํ าก็จะเต่ง เซลล์ท่สูญเสียนํ้ าก็จะแฟบ ใบของพืชที่มกลุ่มเซลล์ชนิดนี้จงกางเมื่อ ี ี ี ึ เซลล์มน้ําและหุบเมือ เซลล์สญเสียนํ้า ี ่ ู ั กลไกการตอบสนองของพืชต่อการเปลียนแปลงของปจจัยภายนอก มีกระบวนการ ่ ดังนี้ การรับสัญญาณ (perception) นักวิทยาศาสตร์ทราบว่าพืชหรือส่วนของพืชมึ ั ความสามารถในการรับสัญญาณการเปลียนแปลงของปจจัยภายนอกทีมากระตุนได้ เช่น ่ ่ ้ มีสารสีททาหน้าทีรบแสง (photo pigment) ทีกระตุนให้เกิดการตอบสนองอย่างมีทศทาง ่ี ํ ่ั ่ ้ ิ ั ของพืชต่อแสง เป็ นต้น แต่สงทีเป็ นปญหา คือ ไม่ทราบว่าสารสีชนิดใดทําหน้าทีน้ี กลไก ิ่ ่ ่ การตอบสนองต่ อ สิ่ง เร้า ชนิ ด อื่น ก็เ ช่น กัน ทัง นี้ เ พราะพืช ไม่ม ีอ วัย วะพิเ ศษที่ทํา หน้ า ที่ ้ ั เฉพาะเจาะจงเพือรับรูและตอบสนองต่อปจจัยกระตุนจําเพาะเหมือนอย่างสัตว์ ่ ้ ้ การส่งสัญญาณ (transduction) คือ กลไกการแปลงและส่งสัญญาณทีรบมาจาก ่ั ั ั ปจจัยกระตุนไปยังเนื้อเยื่อส่วนทีตอบสนองต่อปจจัยชนิดนัน ๆ ของพืช ในช่วงเวลาทีผาน ้ ่ ้ ่ ่ มารวมทังปจจุบน นักวิทยาศาสตร์ได้ศกษารูปแบบของสัญญาณทีพชส่งไปยังเนื้อเยื่อ โดย ้ ั ั ึ ่ ื มุงเน้นทีสญญาณไฟฟ้าและสารเคมี ได้แก่ ฮอร์โมนพืช ่ ่ ั การตอบสนอง (response) คือ การเคลื่อนไหวของพืชวึงเกิดจากการเปลียนแปลง ่ ่ ั ส่วนต่าง ๆ ของพืชทีตอบสนองต่อปจจัยกระตุน ได้แก่ ่ ้ 1) กลไกทีคล้ายคลึงกันในพืชชนิดเดียวกันอาจมีการตอบสนองทีแตกต่างกันได้ ่ ่ ี เช่น กลไกการเปลียนแปลงปริมาตรของเซลล์ทเี่ กิดมาจากการเคลื่อนย้าย K+ ทําให้มการ ่ CU 474 465
54.
เคลือนทีของนํ้าเข้าหรือออกจากเซลล์ มีผลต่อการเปิ ดและปิ
ดของปากใบ ขณะเดียวกันก็ม ี ่ ่ ผลต่อการหุบของใบเมือมีการสัมผัส ่ 2) กลไกทีแตกต่างกันมีผลต่อการตอบสนองทีคล้ายคลึงกันในพืชคนละชนิดหรือ ่ ่ ในเนื้อเยื่อส่วนที่ต่างกันของพืชชนิดเดียวกัน เช่น การโค้งเข้าหาแสงในพืชต่างชนิดกัน อาจเกิดจากกลไกของสารสีทต่างกัน ่ี การตอบสนองของพืชมีการเปลียนแปลงในหลายรูปแบบ ทังด้านชีวเคมี (biochemistry) ่ ้ สรีรวิทยา (physiology) และสัณฐานวิทยา (morphology) แต่การตอบสนองทีสงเกตได้ ่ ั ง่ายทีสด คือ การเคลือนไหว ุ่ ่ การเคลื่อนไหวของพืชที่ตอบสนองต่อสิ่งเร้าโดยมีทิศทางไม่สมพันธ์กบทิศทาง ั ั ของสิ่งเร้า (Nasty หรือ Nastic Movement) ใบ เกล็ด หุ้ ม ตา และดอกของพืช บางชนิ ด มี ก ารตอบสนองแบบ nastic movement คือ การบานหรือกางออกซึ่ง เป็ นการโค้งงอลงเรียกว่า epinasty และมี การหุบ คือ การโค้งงอขึนของกลีบดอกหรือ ้ ใบ เรียกว่า hyponasty การเคลื่อนไหวโดย การโค้งงอของส่วนกลีบดอกและใบมักเกิด จากกลุ่มเซลล์ใน pulvinus บริเวณโคนใบ รูปที่ 10.4.1 กลุ่มเซลล์ใน pulvinus ของพืช โคนก้านใบ หรือโคนใบย่อย แต่ในพืชบาง ชนิดก็เกิดจากการเจริญเติบโตของเซลล์ดานบนกับด้านล่างไม่เท่ากัน เช่น กลีบหรือใบที่ ้ เนื้อเยื่อด้านในเจริญเติบโตเร็วกว่าด้านนอก ทําให้กลีบดอกหรือใบกางออก ในทางตรงกัน ข้ามกันก็จะทําให้เกิดการหุบของกลีบดอกหรือใบ 1. Nyctinasty เป็ นการเคลื่อนไหวโดยแผ่นใบกางออกรับแสงในเวลากลางวัน และ หุบในเวลากลางคืน การบานของดอกในเวลาเช้าและหุบในเวลากลางคืน เช่น ดอกบัว จึง ั มีความสัมพันธ์ระหว่างปจจัยสิงแวดล้อมภายนอก และนาฬิกาชีวภาพภายในของพืช การ ่ หุบของใบพืชเกิดจากกลไกการเคลื่อนที่ของนํ้ าที่ตอบสนองต่อการเปลี่ยนแปลงความ ํ ู เข้มข้นของอิออนภายในเซลล์ เช่น เมือ K+ เคลื่อนออกจากเซลล์ทาให้สญเสียนํ้า ลดความ ่ 466 CU 474
55.
เต่งลง ในขณะที่ K+
เคลื่อนเข้าสู่เซลล์ ทําให้เซลล์ได้รบนํ้ าจึงเต่งขึ้น การเปลี่ยนแปลง ั ความเต่งของเซลล์ ทําให้สวนของพืชเคลือนไหวได้ ่ ่ ั แสงเป็ น ป จ จัย สํา คัญ ที่ก ระตุ้น ให้ เกิด การหุ บ พืช ที่ใ บตอบสนองต่ อ กลางวัน และกลางคืน จะมีแสงเป็ นตัวกระตุ้นให้เซลล์ บางส่ ว นเพิ่ม ความเต่ ง และขยายขนาดใน ขณะที่เซลล์บางส่วนสูญเสียความเต่ง โดยมี รูปที่ 10.4.2 การบานของดอกทีมแสงเป็ นสิงเร้า อิทธิพลของนาฬิกาชีวภาพภายในขอพืชร่วม ่ ี ่ ด้วย ซึงพบว่าการกางและหุบของใบใกล้เคียง ่ กับเวลาเช้าและเย็น แม้พชจะอยูในความมืดต่อเนื่องเป็ นระยะเวลาหนึ่ง ื ่ 2. Hydronasty เป็ นการเคลื่อนไหวของพืชทีตอบสนองโดยการพับหรือม้วนใบต่อ ่ การขาดนํ้ า ซึ่งเป็ นการช่วยลดการคายนํ้ าโดยการลดพื้นที่ของใบในการรับแสง การพับ หรือม้วนของใบหญ้าเกิดจากการสูญเสียความเต่งใน bulliform cell ทีผวใบ ซึงเป็ นเซลล์ ่ ิ ่ พิเศษทีม ี Cuticle น้อยมาก จึงสูญเสียนํ้าได้เร็วกว่าเซลล์ผว (epidermis) อื่น ๆ เมือความ ่ ิ ่ เต่งในเซลล์ลดลง แรงเต่งของเซลล์ทผวใบด้านล่างทําให้ใบพับลง ่ี ิ 3. Thigmonasty เป็ นการเคลื่อนไหวของพืชทีตอบสนองต่อการสัมผัส พบในพืชต่าง ่ ๆ โดยเฉพาะพืชตระกูลถัว เช่น ไมยราพ แม้ได้รบการสัมผัสเพียงใบย่อยใบเดียวอาจทํา ่ ั + ่ ให้ใบหุบได้ทงต้น เกิดจากกลไลการเคลื่อนย้าย K ทําให้เกิดการเปลียนแปลงความเต่งใน ั้ เซลล์ การหุบอย่างรวดเร็วของใบช่วยไล่แมลงทีมารบกวน ่ ใบของต้น กาบหอยแครง (Venus’s-Flytrap) มีข นพิเ ศษที่ไ วต่ อ การสัม ผัส เมื่อ แมลงสัมผัสกับขนจะทําให้เกิดการเปลียนแปลงศักย์ไฟฟ้า (Action potential) เคลื่อนจาก ่ ขนไปที่เนื้อเยื่อของใบกับดัก ซึ่งจะปิ ดลงเพียงครึงวินาทีเท่านัน จากนัน เอนไซม์จะถูก ่ ้ ้ ปล่อยออกมาย่อยแมลงซึงจะเป็ นแหล่งอาหารพวกแร่ธาตุไนโตรเจน และฟอสเฟตของพืช ่ CU 474 467
56.
ก. ข. รูปที่ 4.4.3 การตอบสนองต่อการสัมผัสของใบพืช
ใบไมยราพ (ก) และใบกับดักแมลงของต้นกาบหอยแครง (ข) 4. Thigmomorpho genesis เป็ นการตอบสนองต่ออิทธิพลกระตุนทางกายภาพทีม ี ้ ่ ผลต่อการเจริญเติบโตและพัฒนาของพืช นักวิทยาศาสตร์ทดลองลูบบริเวณลําต้นพืชเบา ๆ แล้วพบว่าพืชลดอัตราการยืดของลําต้นทําให้ลําต้นมีขนาดใหญ่ เตียและแข็งแกร่งมากขึน ้ ้ ั ลมเป็ นปจจัยหนึ่งทีกระตุนให้พชตอบสนองโดยมีลําต้นเตียลง เช่น alfalfa ทีเจริญเติบโต ่ ้ ื ้ ่ ในบริเวณทีมลมแรงปานกลาง การโยกไหวของลําต้นทําให้มความสูงลดลงถึง 36% เมื่อ ่ ี ี เทียบกับต้นทีถูกจํากัดการเคลือนไหวของลําต้น ่ ่ การเคลื่อนไหวของพืชที่ตอบสนองอย่างมีทิศทางต่อทิศทางของปัจจัยสิ่งเร้า (Tropic movement หรือ Tropism) Tropism เป็ นการเคลื่อนไหวของพืชโดยมีทศทางเกียวข้องสัมพันธ์กบทิศทางของ ิ ่ ั สิงเร้า การเคลื่อนไหวนี้ทําให้สวนของพืชโค้งเข้าหา (positive) หรือหนี (negative) สิงเร้า ่ ่ ่ ทีมากระตุน ซึงเกิดจากสิงเร้าไปกระตุนฮอร์โมนแล้วฮอร์โมนไปกระตุนให้สวนต่าง ๆ เกิด ่ ้ ่ ่ ้ ้ ่ การเจริญเติบโตไม่เท่ากัน ทําให้เกิดการโค้งงอของส่วนนันของพืช Tropism จําแนกตาม ้ ชนิดของสิงเร้าได้ดงนี้ ่ ั 468 CU 474
57.
1. Phototropism เป็
นการตอบสนอง อย่างมีทศทางต่อแสง ปลายยอดมีการเติบโต ิ โดยเคลื่ อ นไหวเข้ า หาแสงเป็ น positive phototropism เนื่องจากปลายยอดด้านไม่ได้ รั บ แสงมี อ ั ต ราการยื ด ขยายของเซลล์ มากกว่าด้านที่ได้รบแสง ทําให้เกิดการโค้ง ั ง อ เ ข้ า ห า แ ส ง ส่ ว น ก า ร ง อ ก แ ล ะ ก า ร เจริญ เติบ โตของรากจะเคลื่อ นไหวเบนหนี รูปที่ 10.4.4 การตอบสนองของพืชต่อแสงสว่าง แสงเป็ น negative phototropism เกิดจาก ปลายรากด้านรับแสงมีการยืดขยายของเซลล์มากกว่าด้านไม่ได้รบแสง จึงทําให้เกิดการ ั โค้งออกจากแสง 2. Gravitropism เป็ นการตอบสนองอย่างมีทศทางต่อแรงโน้มถ่วงของโลก การงอกและ ิ เจริญเติบโตของรากจะมีทศทางเข้าหาแรงดึงดูดของโลกเป็ น positive gravitropism ส่วน ิ การงอกและเจริญ เติบ โตของยอดและลํ า ต้ น จะมีทิศ ทางหนี แ รงดึง ดู ด ของโลกเป็ น negative gravitropism 3. Hydrotropism เป็ นการตอบสนองอย่างมีทศทางต่อความชืน เมือรากมีอายุมาก ิ ้ ่ ขึนจะมีการตอบสนองต่อแสง ความชืน และการสัมผัส การตอบสนองต่อความชืน รากจะ ้ ้ ้ เจริญเติบโตเข้าสู่บริเวณทีมศกย์ของนํ้ ามาก ทําให้บางครังเป็ นไปในทิศทางเดียวกับแรง ่ ี ั ้ โน้มถ่วงของโลก รากจะเติบโตเข้าหานํ้าหรือความชืนส่วน hypcotyl จะเติบโตหนีน้ําหรือ ้ ความชืน ้ 4. Thermotropism เป็ นการเคลื่อนไหวของพืชโดยมีอุณหภูมเิ ป็ นสิงเร้า เช่น การ ่ ออกดอก และการงอกของเมล็ดพืชบางชนิดในอุณหภูมต่า เป็ นต้น ิํ 5. Chemotropism เป็ นการเคลือนไหวของพืชโดยมีสารเคมีเป็ นสิงเร้า เช่น รากจะ ่ ่ งอกเข้าหา peptone และ asparagines ซึงมีอยูในอาหาร ละอองเกสรทีตกลงบนยอดเกสร ่ ่ ่ ตัวเมีย จะงอกหลอดละอองเรณู (pollen tube) เข้าหากลูโคส หรือกรดมาลิก (malic acid) ในรังไข่ 6. Thigmotropism เป็ นการเคลื่อนไหวของพืช โดยมีการสัมผัสเป็ นสิงเร้า พืชบาง ่ ชนิดมีมอเกาะ (tendril) ทําหน้ าที่ยดเกาะเพื่อพยุงลําต้น พืชหลายชนิดไม่มมอเกาะแต่ ื ึ ี ื เจริญเติบโตโดยใช้ลําต้นเลื้อยพันกับสิงที่ยดเกาะได้ การตอบสนองแบบนี้มประโยชน์ ่ ึ ี CU 474 469
58.
สําหรับพืชมากเพราะช่วยให้พชสามารถแข่งขันกับพืชชนิดอื่น ๆ ในการรับแสง
รากของ ื พืชสามารถตอบสนองอย่างมีทศทางต่อการสัมผัสได้ดวย ิ ้ การเคลื่อนไหวที่เกิดจากปัจจัยภายในต้นพืช ั การเคลื่อนไหวของพืชอันเนื่องมาจากป จจัยหรือสิงเร้าภายใน คือ ฮอร์โมนพืช ่ เรียกว่า การเคลือนไหวอัตโนวัติ (Autonomic Movement) ได้แก่ ่ 1. Nutation หรือ Nutational Movement เป็ นการเคลื่อนไหวแบบหมุนส่าย เนื่องจาก การเจริญเติบโตของยอดไม่เท่ากันทําให้ปลายยอดเอนไปมาเมือพืชเจริญเติบโตขึนทีละน้อย ่ ้ 2. Spiral Movement เป็ นการเคลื่อนไหวทีบดเป็ นเกลียวของยอดพืชเมื่อพืช ่ ิ เจริญเติบโตขึน แต่พชบางชนิดปลายยอดจะบิดเป็ นเกลียวรอบแกนตังตรงขึนไปซึงสังเกต ้ ื ้ ้ ่ ได้ย าก ซึ่ง ความเป็ น จริง ซีก ซ้า ยและขวาจะเจริญ ไม่เ ท่ า กัน จึง มีก ารบิด ซ้า ยขวาบ้า ง เล็กน้อย ซึงมักเรียกว่า Circumnutation ่ 3. Twining movement เป็ นการ เคลื่ อ นไหวที่ ส่ ว นยอดของพื ช บิ ด เป็ น เกลีย ว โค้ ง อ้ อ มพัน หลัก ขึ้น ไปเรื่อ ย ๆ เพือพยุงลําต้นทีอ่อนและมีขนาดเล็ก ่ ่ 4. Nodding Movement เป็ นการ รูปที่ 10.4.5 การเจริญของไม้เลือยพันหลักเป็ น ้ เคลื่อนไหวของพืชเพื่อชูสวนใบเลียงขึนมา ่ ้ ้ เกลียวเพือพยุงลําต้น ่ เหนือดินขณะเมล็ดงอก เช่น เมล็ดถัวบาง ่ ชนิด ขณะเมล็ดงอกมีการสร้างเอธิลน ทีทา ี ่ ํ ให้ ลํ า ต้ น ใต้ ใ บเลี้ย ง (hypocotyl) โค้ ง งอ โผล่ พ้น ดิน ขึ้น มาพร้อ มกับ การโยกไปมา รูปที่ 10.4.6 การงอกของเมล็ดถัว ่ ช่วยให้ต้นกล้าดันดินโผล่ส่วนของใบเลี้ยง ขึนมาเหนือดิน ้ 470 CU 474
59.
รูปที่ 10.4.7 การเจริญของต้นลิลลี่ 5.
Contraction Movement เป็ นการเคลื่อนไหว ของพืชทีเกิดจากส่วนของพืชหดตัวแล้วไปมีผลต่อ ่ การดึงส่วนใดส่วนหนึ่งของพืช เช่น ต้นลิลลี่ ทีเพาะ ่ เมล็ดบนผิวดินมีรากหดตัว (Contractile root) ช่วย ดึงให้ลําต้นเคลื่อนตัวลงไปในดินพร้อมกับสร้างหัว ใต้ผวดินการเคลื่อนไหวแบบอัตโนวัตของพืชนี้จะ ิ ิ ไม่ เ กี่ ย วข้ อ งกั บ สิ่ ง แวดล้ อ มแม้ ว่ า สิ่ ง แวดล้ อ ม ภายนอกจะเปลี่ย นไป เช่ น เวลากลางวัน หรือ กลางคืนแสงมากหรือแสงน้อย การเคลื่อนไหวของ พืชก็ยงคงดําเนินต่อไป ั การตอบสนองของพืชต่อสารควบคุมการเจริญเติบโต ฮอร์โมนพืช (Plant hormone or Phytohormone) ฮอร์โมนพืชเป็ นสารควบคุมการเจริญเติบโตของพืช (Plant growth regulator) ั ั และเป็ นปจจัยภายในของพืชซึงต้องขึนอยู่กบปจจัยพันธุกรรม (Genetic factor) และปจจัย ่ ้ ั ั สิงแวดล้อม (Environment factor) เป็ นตัวควบคุม แบ่งเป็ น 5 กลุ่ม คือ Auxin Gibberellin ่ ั ั Cytokinin Ethylene และ Abscisin แต่ในปจจุบนนักวิทยาศาสตร์เชือว่าสารกลุ่ม Steroid ่ ในพืช ก็เ ป็ น ฮอร์โ มนชนิ ด หนึ่ งที่ม ีผ ลต่ อ การเจริญ เปลี่ย นแปลงรูป ร่า งที่ช ก นํ า ด้ว ยแสง ั นอกจากนี้ยงพบสารอื่น ๆ เช่น jasmonic acid, Salicylic acid และ Systemin ซึงเป็ น ั ่ โปรตีน ก็อาจเป็ นฮอร์โมนพืชด้วยเช่นกัน ออกซิ น (Auxin) ออกซินเป็ นฮอร์โมนพืชชนิดแรกที่นักวิทยาศาสตร์ค้นพบ ชนิดทีรูจกกันดี คือ 3 ่้ ั Indoleacetic acid (IAA) ซึงพบโดย Charles Dawin ในปี ค.ศ. 1880 โดยเขาสังเกตเห็น ่ ว่าทีสวนปลายของแผ่นหุมใบเกิดใหม่ (coleoptile) ของต้นกล้า canary grass จะโค้งเข้า ่่ ้ หาแสงเสมอ เขาทําการทดลองโดยใช้กระดาษตะกัวปิ ดปลายยอดของต้นกล้าไม่ให้ได้รบ ่ ั แสง พบว่า ส่วนเนื้อเยื่อทีอยู่ต่ํากว่าปลายยอดลงมามีการเจริญเติบโตของเนื้อเยื่อต่างกัน ่ และทําให้coleoptiles เอนเข้า หาแสงและจากการทดลองโดยหุ้มส่วนที่อยู่ต่ํากว่า coleoptiles ด้วยกระดาษตะกัวโดยปล่อยให้ปลายยอดได้รบแสง พบว่า ส่วนปลายนันโค้ง ่ ั ้ CU 474 471
60.
เข้าหาแสงได้ ต่อมาเขาได้ทําการทดลองใหม่โดยการตัดปลาย coleoptiles
ออกประมาณ 2.5-4.0 มิลลิเมตร ผลการทดลองพบว่าปลายยอดไม่โค้งเข้าหาแสง Darwin สรุปการ ทดลองว่า ทีปลายยอดมีการสร้างสารสือสัญญาณ (Signal) เคลื่อนลงมาทีสวนเจริญเติบโต ่ ่ ่่ และทําให้สวนทีไม่ถูกแสงเจริญเร็วกว่าส่วนทีได้รบแสง ผลงานนี้ลงพิมพ์ใน The power of ่ ่ ่ ั Movement in Plants’ ซึงเป็ นสิงพิมพ์ทมความสําคัญทางวิชาการในปลายทศวรรษที่ 18 ่ ่ ่ี ี ก. ข. รูปที่ 10.4.8 การทดลองของ Charles Dawin (ก) และขันของการงอกของเมล็ดโอ๊ต (ข) ้ Boysen-Jensen (1913) ทดลองตัดปลาย coleoptile ของต้นกล้า ข้าวโอ๊ต แล้ว วางก้อนวุน (gelatin) ลงบน coleoptile ส่วนทีเหลือแล้วนํ าส่วนทีตดออกวางบนก้อนวุน ้ ่ ่ ั ้ ปรากฏว่ายอดข้าวโอ๊ตโค้งเข้าหาแสง นอกจากนี้ยงทดลองเฉือน coleoptile ด้านทีไม่ได้รบ ั ่ ั แสงตามขวาง แล้วสอดแผ่นไมก้าเข้าไปคันเอาไว้ พบว่ายอดข้าวไม่โค้งเข้าหาแสง แต่ตน ่ ้ ทีเฉือนและสอดไมก้าทางด้านทีได้รบแสง ปรากฏว่ายอดข้าวโค้งเข้าหาแสง ่ ่ ั จากการทดลองของ Boysen-Jensen แสดงว่ามีการลําเลียงสารจากปลาย coleoptile ลงมาทางด้านล่าง สามารถแพร่ผ่านก้อนวุ้น แต่ไม่สามารถผ่านแผ่นไมก้า เมื่อด้านทีไม่ได้ ่ รับแสงมีการเคลือนย้ายสารลงมาได้ จะกระตุน ่ ้ เนื้อเยือ coleoptile เติบโตเข้าหาแสงได้ ่ Paal (1919) ทําการทดลองโดยตัด รูปที่ 10.4.9 การทดลองของ Boysen-Jensen ปลาย coleoptile ของกล้า ข้า วโอ๊ ต นํ า ไปวาง 472 CU 474
61.
บน coleoptile ที่เหลือในตําแหน่
งที่เลื่อนไปอยู่ขางที่ไม่ได้รบแสง พบว่า ปลายยอดเอน ้ ั โค้งเข้าหาแสง เขาสรุปว่าสือสัญญาณกระตุนการเจริญเติบโตนี้น่าจะเป็ นสารเคมี ่ ้ Went (1926) นําแนวคิดทีสารซึมผ่านก้อนวุนได้ ก็น่าจะเก็บสะสมไว้ได้ เขาทําการ ่ ้ ทดลองโดยตัดปลาย coleoptile ของกล้าข้าวโอ๊ตมาวางบนก้อนวุนเป็ นเวลา 1-2 ชัวโมง ้ ่ แล้วนําก้อนวุนไปวางบน coleoptile ส่วนทีเหลือ ปรากฏว่า coleoptile เติบโตได้ตามปกติ ้ ่ และเมื่อทดลองนํ าก้อนวุนไปวางบนด้านใดด้านหนึ่งได้ แสดงว่ามีสารแพร่จากก้อนวุนไป ้ ้ ทําให้เนื้อเยื่อ coleoptile ขยายตัว ต่อมา F. Kogl และคนอื่น ๆ ได้ตงชือสารนี้วา auxin ั้ ่ ่ ซึงมีรากศัพท์มาจากภาษากรีก คือ Auxen ซึงมีความหมายว่า การเพิมขึนหรือเติบโต (to ่ ่ ่ ้ increase or togrow) ก. ข. รูปที่ 10.4.10 การทดลองของ Went (ก) และทดสอบโดย Avena (ข) ปี ค.ศ. 1930 F. Kogl และ A. J. Haagen-Smit ค้นพบ ออกซินในพืช คือ Indoleั 3-acetic acid (IAA) จากการแยกสารบางอย่างจากนํ้าปสสาวะของคน ซึงนําไปทดลองกับ ่ coleoptile แล้วได้ผลเช่นเดียวกับสารที่ Went แยกออกมาจาก coleoptile จึงเชือว่าสารใน ่ coleoptile ก็คอ IAA การศึกษาในเวลาต่อมาพบว่า IAA เป็ น auxin ทีพบมากทีสดในพืช ื ่ ุ่ และมีฤทธิทางชีววิทยา ต่อมานักวิทยาศาสตร์ได้สงเคราะห์ auxin ขึนมาอีกหลายชนิด ั ้ ์ ได้แก่ 3-Indole butyric acid (IBA) 2,4 Dichloro plonic acid (IPA) α - Naphthalene acetic acid (NAA) β - Naphthoxy acetic acid (NOA) 2,4 Dichlorophenoxy acetic acid (2,4-D) 2,4,5-Trichlorophenoxy acetic acid (2,4,5-T) และ 2-MeHuyl 4chlorophenoxy acetic acid (MCPA) CU 474 473
62.
คุณสมบัติของออกซิ น 1. ออกซินสร้างที่ยอดและลําเลียงลงล่างเสมอ
มีโพลาริตี (polarity) ซึ่งสามารถ ตรวจสอบได้จากการทดลองตัดปลาย coleoptile ของพืชมาวางบนก้อนวุนเป็ นระยะเวลา ้ หนึ่ง แล้วนํ าก้อนวุนทีมออกซินไปวางบนท่อน coleoptile ทีตดออกมาจากตอส่วนทีเหลือ ้ ่ ี ่ ั ่ นํ าไปวางบนก้อนวุนเปล่าอีกก้อนหนึ่ง ปรากฏว่าออกซินจากวุนก้อนบนจะแพร่ผ่านท่อน ้ ้ coleoptile ลงมาทีวนก้อนล่างจนทําให้มออกซินมากกว่าก้อนบน ่ ุ้ ี รูปที่ 10.4.11 การทดลอง polarity ของออกซิน ต่อมาพลิกท่อน coleoptile กลับเอาด้านล่างขึนข้างบน พบว่า ออกซินจากวุนก้อน ้ ้ บนไม่แพร่ลงมาทีวนก้อนล่าง ่ ุ้ Polority นี้พบมีเฉพาะในลําต้น ใบ ก้าน เท่านัน ไม่พบในรากซึงออกซินสามารถ ้ ่ ลําเลียงได้ทงสองทาง ั้ 2. ออกซินไวต่อแสง ทียอดพืชมีการสร้างออกซินจึงมีปริมาณมาก เมื่อได้รบแสง ่ ั สว่าง ออกซินจะถูกลําเลียงหนีแสง ดังนันด้านทีไม่ได้รบแสงจึงมีออกซินปริมาณมากกว่า ้ ่ ั ด้านทีได้รบแสง นอกจากนี้แสงยังเป็ นตัวทําลายออกซินโดยทําให้ออกซินเปลียนเป็ นสาร ่ ั ่ ชนิดอื่น ซึงนับว่าเป็ นผลดีของพืช ทังนี้เนื่องจากฮอร์โมนตามธรรมชาติมฤทธิ ์รุนแรงมาก ่ ้ ี ถ้า พืช สร้า งและสะสมมากอาจทํ า ให้พ ืช ได้ร บ อัน ตราย พืช จึง มีก ลไกการรัก ษาระดับ ั ฮอร์โมนให้พอดี โดยแสงจะทําให้เกิด oxidation เปลียน IAA ไปเป็ น Indolealdehyde ซึง ่ ่ เป็ นสารอินทรียทไม่ทางานจึงไม่ทาให้เกิดการยืดขยายตัวของเนื้อเยือ ์ ่ี ํ ํ ่ 474 CU 474
63.
IAA indoleacetic acid oxidase
indolealdehyde + H2O + CO2 Light (inactive) 3. ควบคุมการเจริญเติบโตของลําต้น ตา ใบ และราก ทีแตกต่างกัน คือ ทีระดับ ่ ่ ความเข้มข้นของออกซินทีพอเหมาะกับการเจริญเติบโตดีทสดของลําต้น จะเข้มข้นเกินไป ่ ่ี ุ สําหรับการเจริญของตาและใบ จึงเป็ นความเข้มข้นทียบยังการเจริญเติบโตของตาและใบ ่ ั ้ และการเจริญเติบโตของรากต้องการความเข้มข้นของออกซินทีต่ากว่าทีตาและใบต้องการ ่ ํ ่ มาก รูปที่ 10.4.12 ความเข้มข้นของออกซินทีมผลต่อการเจริญของลําต้นและราก ่ ี พืชทุกชนิดสามารถสร้างออกซินได้ทงในลําต้น ราก ใบ ก้าน ดอก ผล เมล็ด และ ั้ แม้แต่ในละอองเกสร ในปริมาณทีแตกต่างกัน ปลายยอด และปลาย coleoptile ในต้นอ่อน ่ ของพืชใบเลียงเดียวมีการสร้างมากทีสด รองลงมาคือ ส่วนทียงอ่อน เช่น ใบอ่อน รากอ่อน ้ ่ ุ่ ่ ั จะมีการสร้างมากกว่า ใบและรากทีมอายุมาก สารทีใช้สร้าง IAA คือ Tryptophan และ ่ ี ่ เอนไซม์หลายชนิดทีตาและส่วนอื่น ๆ ของพืช นอกจากนี้ยงมีแร่ธาตุทจําเป็ นต่อการสร้าง ่ ั ่ี Tryptophan คือ สังกะสีรวมอยู่ดวย IAA ในเซลล์พชมีทงทีอยู่ในไซโตซอลโดยจับอยู่กบ ้ ื ั้ ่ ั สารอื่น และเป็ น IAA อิสระ อยู่ในคลอโรพลาสต์ ซึงจะถูกควบคุมปริมาณด้วย IAA ในไซ ่ โตซอลทําให้มความสมดุล ี CU 474 475
64.
อิทธิพลของออกซินต่อกระบวนการสรีรวิทยาของพืช 1. ควบคุมทิศทางการเจริญเติบโตและพัฒนาของพืช โดยตอบสนองอย่างมีทศทาง ิ ั ั ต่อปจจัยภายนอก
(Plant tropism) ในสองปจจัยคือ แสง และแรงโน้มถ่วง Phototropism เป็ นปรากฏการณ์ทพบในลําต้นของพืชทุกชนิดกับในรากของพืช ่ี หลายชนิด เป็ นกระบวนการทีมความสําคัญต่อการดํารงชีวตและเผ่าพันธุ์ของพืช เพราะ ่ ี ิ ทําให้ใบของพืชเติบโตในทิศทางทีจะได้รบแสงเพื่อทําการสังเคราะห์ดวยแสง ในการสร้าง ่ ั ้ อาหาร ส่วน gravitropism เป็ นกระบวนการทีทาให้รากเจริญเติบโตในทิศทางทีลงสูพนดิน ่ ํ ่ ่ ้ื และลําต้นพืชเติบโตในทิศทางพุงตรงไปในอากาศ ซึงมีความสําคัญต่อการอยูรอดของพืช ่ ่ ่ 2. ควบคุมการเจริญเติบโตของเนื้อเยื่อและอวัยวะ ได้แก่ ตา แคมเบียม ราก และ ลําต้น โดยออกซินไปกระตุนเซลล์ให้มการสร้างสารทีเป็ นองค์ประกอบของผนังเซลล์สะสม ้ ี ่ ไว้มากทีปลายเซลล์ทง 2 ข้าง ทําให้เซลล์ยดขยายยาวขึน (cell elongation) ่ ั้ ื ้ 3. ควบคุมการแบ่งเซลล์แคมเบียมโดยออกซินเร่งการแบ่งตัวของเซลล์แคมเบียม ในพืชใบเลียงคู่ ทําให้ลาต้นขยายขนาดใหญ่ขน ้ ํ ้ึ 4. ควบคุมการเจริญของตาข้าง (Lateral bud) โดยตายอด (terminal bud) สร้าง ออกซินลําเลียงลงมาด้านล่าง ออกซินปริมาณมากจะยับยังการเติบโตของตาข้าง เรียกว่า ้ การข่มของตายอด (Apical dominance) ก. ข. รูปที่ 10.4.13 การทดลองผลของออกซินทียอดยับยังการเจริญของตาข้าง (ก) ่ ้ และออกซินจากยอดข่มตาข้างของมันฝรัง่ (ข) 476 CU 474
65.
5. ควบคุมการงอกรากของกิงตัดหรือกิงตอน การตอนกิงทําโดยการลอกเปลือกไม้ ่ ่ ่ ออก
ทําให้ออกซินที่ลําเลียงลงมาจากยอดจะมาสะสมอยู่ท่บริเวณรอยแผลด้านบน และ ี เซลล์ของเนื้อเยือเจริญสามารถแบ่งเซลล์เปลียนแปลงเป็ นรากได้ ่ ่ 6. ควบคุมการมีดอกของพืช พืชทีเติบโตจนใกล้จะมีดอกถ้าได้รบออกซิน จะทําให้ ่ ั ยืดระยะเวลาการออกดอก ทําให้ออกดอกช้า ั 7. ควบคุมการเจริญเติบโตของผล การเจริญเติบโตของผลต้องอาศัยปจจัยการ เติบโต (Growth factor) จากต้นอ่อน (embryo) และอาหารเลียงต้นอ่อน (endosperm) ้ ภายในเมล็ด ถ้าโอวุล (ovule) ในรังไข่ไม่ถูกผสมเกสร โอวุลจะไม่เจริญเป็ นเมล็ดและรังไข่ ไม่เจริญไปเป็ นผลแต่เราสามารถทําให้ผลไม้บางชนิดเจริญเติบโตโดยไม่มเี มล็ดได้ เช่น สต อรเบอรี่ มะเขือเทศ องุน แตงโม บวบ และมะเดื่อ โดยใช้ออกซิน เช่น IAA, 2, 4-D หรือ ่ gibberellin ทีความเข้มข้นพอเหมาะ พ่นหรือป้ายทีดานเกสรตัวเมียซึงยังไม่ได้รบการผสม ่ ่ ้ ่ ั รังไข่จะเจริญไปเป็ นผลทีไร้เมล็ดเรียกผลไม้ชนิดนี้วา Parthenocarpic fruit ่ ่ 8. ควบคุมการร่วงของใบ ดอก และผล กระบวนการร่วงของใบ ดอก และผล (abscission) เป็ นการเปลี่ยนแปลงทางสรีรวิทยาซึ่งเกิดในบริเวณก่อการร่วง (abscission zone) ที่ใกล้กบโคน ก้าน ใบ ซึ่งแตกต่างจากการเสื่อมสลาย (senescence) ที่หมายถึง ั การเปลียนแปลงทีนําไปสูความตายของพืชทังต้นหรือส่วนหนึ่งส่วนใด การหลุดร่วงของใบ ่ ่ ่ ้ พืชเริมจากการเปลียนแปลง (differentiation) ของชันเนื้อเยื่อก่อการร่วง โดยผนังเซลล์จะ ่ ่ ้ ถูกย่อยด้วยเอนไซม์ cellulose และ pectinase ทําให้เซลล์อ่อนตัวและแยกออกจากกัน ปริมาณออกซินมีมากในใบอ่อน และจะค่อย ๆ ลดลงในใบแก่จนน้อยทีสด เมือใบเริมเสือม ุ่ ่ ่ ่ เข้าสูความตาย IAA จะช่วยชะลอการหลุดร่วงของใบในระยะแรก แต่จะเร่งในระยะหลังทํา ่ ให้ใบหลุดร่วงเร็วขึน ทีเป็ นเช่นนี้เกิดจากการทีใบมีการสร้างเอธิลนมากขึนเมือปริมาณออก ้ ่ ่ ี ้ ่ ซิน ลดลง เอธิลีน เป็ น สารที่ช่ ว ยในการหลุ ด ร่ ว งของใบ โดยทํ า ให้เ นื้ อ เยื่อ ก่ อ การร่ ว ง (abscission layer) ทีโคนก้านเจริญเร็วเกินไปจนเกิดการแยกตัวของเซลล์ตามแนวนัน ่ ้ ส่วนการร่วงของส่วนต่าง ๆ ของพืช เป็ นผลทางอ้อมของออกซินโดยออกซินไปกระตุนให้ ้ เกิดการสร้าง abscisin (abscisic acid) CU 474 477
66.
รูปที่ 10.4.14 ออกซินควบคุมการร่วงของใบ 9.
การใช้ออกซินสังเคราะห์เป็ นยาฆ่าวัชพืช (Auxin as herbicide) การใช้ออกซิน ทีมความเข้มข้นสูงกว่าธรรมชาติมาก ๆ กับวัชพืช ทําให้มการเจริญทีผดปกติ เช่น เร่งให้ม ี ่ ี ี ่ ิ การแบ่งเซลล์มาก แต่ไม่มการยืดตัว เนื้อเยื่อถูกทําลาย เช่น phloem ถูกทําลายทําให้ ี ลําเลียงอาหารไม่ได้ มีเมแทบอลิซึมผิดปกติ โดยเฉพาะการหายใจที่รวดเร็วเผาผลาญ อาหารทีสะสมไว้ให้หมดไปทําให้พชตาย ่ ื จิ บเบอเรลลิ น (Gibberellin) จิบเบอเรลลิน เป็ นฮอร์โมนพืชทีพบโดยนักพฤกษศาสตร์ชาวญีปุนชือ E. Kurosawa ที่ ่ ่ ่ ่ พบว่า รา Gibberella Fujikurol ทําให้เกิดโรค foolish seedling หรือ bakanae คือต้นข้าว เติบโตสูงผิดปกติ และไม่สร้างเมล็ด เนื่องจากราชนิดนี้ ปล่อยสารชนิดหนึ่งออกมา ซึ่ง ภายหลังเมือมีการแยกสารชนิดนี้ได้จากเชือราจึงเรียกว่า Gibberellin ตามชือของเชือราใน ่ ้ ่ ้ เวลาต่อมานักวิทยาศาสตร์สามารถสกัดสารทีคล้ายคลึงกับ Gibberellin ได้จากพืชหลาย ่ ั ั ชนิด และใช้ระบบการตังชือด้วยเลขหมาย เป็ น Gibberellin Ax ตามลําดับการค้นพบ ปจจุบน ้ ่ พบมากกว่า 40 ชนิด เช่น GA1, GA2, GA3, … แม้จะพบ Gibberellin มากชนิดในพืช แต่ก็ มีเพียงไม่กชนิดทีสารมารถมีกจกรรมทางชีววิทยา และจัดอยู่ในกลุ่มฮอร์โมน ในพืชชันสูง ่ี ่ ิ ้ Gibberellin สร้างมาจากใบอ่อนและผลทียงไม่แก่ ่ ั อิ ทธิ พลของ Gibberellin ต่อการเจริ ญเติ บโตและพัฒนาของพืช 1. กระตุ้นการเจริญเติบโตของลําต้นในพืชแคระ ให้มความสูงเท่ากับพืชปกติ แต่ ี ขนาดความหนาของลําต้นลดลง ใบเล็กและมีสเี ขียวจาง 2. กระตุ้นการยืดตัวและการแบ่งเซลล์ของลําต้น ทําให้ลําต้นสูงขึน แต่ยบยังการ ้ ั ้ ยืดตัว และการเกิดรากใหม่ 478 CU 474
67.
3. กระตุ้นการแบ่งเซลล์แคมเบียมทําให้เกิดโฟลเอมใหม่ ๆ
(ออกซินกระตุ้นการ แบ่งเซลล์แคมเบียมเพือการสร้างไซเลมใหม่ ๆ) ่ 4. กระตุนให้พชวันยาว (long-day plant) ให้ออกดอกเร็วขึน เช่น ผักกาดหอม ้ ื ้ และกะหลํ่าปลี 5. กระตุนการงอกของเมล็ดและการเจริญเติบโตของต้นอ่อนธัญพืชบางชนิด เช่น ้ ข้าวบาร์เลย์ ข้าวสาลี และข้าวโอ๊ต 6. ยับยังการเจริญเติบโตของตาดอก (flower bud) และตาลําต้น (vegetative bud) ้ และลดระยะพักตัวของเมล็ด พืชบางชนิดให้สนลงทําให้เมล็ดงอกได้เร็วขึน ั้ ้ 7. ช่วยให้มการติดผลหลังการถ่ายละอองเรณู และช่วยในการเจริญเติบโตของผล ี พืชบางชนิด เช่น แอปเปิ้ล การใช้ประโยชน์ จากจิ บเบอเรลลิ น เกษตรกรใช้ประโยชน์จากจิบเบอเรลลินในด้านต่าง ๆ ได้แก่ การผลิตผลไม้ เช่น การใช้จบเบอเรลลินช่วยให้ก้านผลองุ่นไม่มเมล็ดยืดยาวขึนทําให้ผลไม่เบียดกันมาก องุ่น ิ ี ้ แต่ละผลเติบโตมีขนาดใหญ่ ใช้จบเบอเรลลินช่วยเร่งการงอกของเมล็ดข้าวบาร์เลย์ในการ ิ ทําข้าวมอลท์ในอุตสาหกรรมผลิตเบียร์ ใช้จบเบอเรลลินในการทําให้ออยยืดขยายตัวของ ิ ้ ปล้องในช่วงฤดูหนาว ทําให้ได้ผลผลิตอ้อยเพิมขึน ใช้ในการปรับปรุงพันธุ์พชโดยการลด ่ ้ ื ช่วงระยะเวลาการเจริญก่อนการสืบพันธุ์ (jurenile) ให้สนลง เช่น กระตุ้นให้ต้นบีท ต้น ั้ กะหลํ่าออกดอกได้ ทําให้ผลิตเมล็ดพันธุ์ได้ หรือปรับปรุงพันธุ์พชให้มลกษณะเตี้ย เพื่อ ื ี ั เพิมผลผลิต และป้องกันการล้มของต้นในพวกธัญพืช รวมถึงการใช้สารยับยังการสร้างจิบ ่ ้ เบอเรลลิน เพือการผลิตไม้ดอกไม้ประดับให้มตนเตียเป็ นพุมสวยงาม ่ ี ้ ้ ่ ไซโตไคนิ นหรือไฟโตไคนิ น (cytokinin หรือ phytokinin) ไซโตไคนินเป็ นฮอร์โมนพืชทีควบคุมการแบ่งเซลล์ของพืช ไคเนติน (Kinetin) เป็ น ่ ไซโตไคนินชนิดแรกทีคนพบในธรรมชาติเมื่อปี ค.ศ. 1955 มีมากในนํ้ามะพร้าว และซีตน ่ ้ ิ (zeatin) เป็ นไซโตไคนินทีมมากทีสุดในธรรมชาติโดยเป็ นองค์ประกอบในเอนโดสเปอร์ม ่ ี ่ ของข้าวโพด ไซโตไคนินในธรรมชาติอยู่ในรูปโมเลกุลอิสระมีอยู่ในพืชดอกทุกชนิด มีมาก บริเ วณปลายราก เนื้ อ เยื่อ เจริญ ปลายยอด เมล็ด ที่กํ า ลัง เจริญ เติบ โต และแคมเบีย ม นอกจากนี้ยงพบในสาหร่ายไดอะตอม มอส เฟิ รน สน และในแบคทีเรีย จุลนทรียบางชนิด ั ์ ิ ์ สามารถควบคุมให้เซลล์พชสร้างไซโตไคนินได้ เมื่อเข้าไปอยู่ในพืชจะชักนําให้เนื้อเยื่อพืช ื CU 474 479
68.
แบ่งเซลล์ ในบางกรณีสามารถพัฒนาโครงสร้างพิเศษได้ เช่น
mycorrhizae ที่อาศัย ร่วมกับพืชหลายชนิด สามารถควบคุมให้พชสร้างปมรากได้ ื อิ ทธิ พลของไซโตไคนิ นต่อการเจริ ญเติ บโตและพัฒนาของพืช ไซโตไคนินมีบทบาทต่อกระบวนการสรีรวิทยาและการเจริญเติบโตพัฒนาของพืช โดยทํางานร่วมกับออกซิน ช่วยเร่งให้พชมีการแบ่งเซลล์อย่างรวดเร็ว ในขณะเดียวกันก็ม ี ื การทํางานตรงข้ามกับออกซินในบางเรือง เช่น กระตุนการเจริญของตาข้าง (Lateral bud) ่ ้ จากการศึกษาการทํางานของไซโตไคนินในพืชจํา ลองพันธุ์ สามารถสร้า งรากได้ และ พัฒนาในหลายเรือง เช่น ่ 1. ตายอดสร้างใบจํานวนมาก 2. ใบมีคลอโรฟิลล์มสเี ขียวมากกว่าปกติ ี 3. ใบและก้านใบ อาจสร้างต้นพิเศษ (adventitious shoots) ได้ แม้ใบและก้านใบ จะไม่มบาดแผล ี 4. ใบชะลอการพัฒนาทีนําไปสูการตาย (Leaf Senescence) ่ ่ 5. การข่มของตายอด (Apical dominance) ลดลงมาก 6. พืชสายพันธุทสร้างไซโตไคนินมากกว่าปกติจะชะงักการเจริญเติบโตและมีปล้อง ์ ่ี สัน ้ 7. ทําให้กงชําออกรากน้อยและรากเจริญเติบโตช้ากว่าปกติ ิ่ แอบไซซิ น หรือดอร์มิน (Abscisin or Dormin) แอบไซซิน เป็ นกรดแอบไซสิค (Abscisic acid หรือ ABA) เป็ นฮอร์โมนพืชทีมมาก ่ ี ตามลําต้น ไรโซม (Rhizome) ตา เมล็ด และผลไม้ทุกระยะ อิ ทธิ พลของแอบไซซิ นต่อการเจริ ญเติ บโตและพัฒนาของพืช หน้าทีสวนใหญ่ของแอบไซซินเป็ นสารยับยังการเจริญเติบโต (growth inhibitor) ่่ ้ ดังนี้ 1. ทําให้การเจริญเติบโตของพืชช้าลง แม้จะทํางานร่วมกับจิบเบอเรลลิน 2. ทําให้เมล็ดและตามีระยะการพักตัวยาวขึนจึงมีชอว่า “Dormin” ้ ่ื 3. ยับยังการทํางานของเอนไซม์อะไมเลส ( α - anylase) ในเอนโดสเปอร์มของ ้ เมล็ดธัญพืช 4. กระตุนการแก่ของใบทีพฒนาไปสูความตาย (leaf senescence) ้ ่ ั ่ 5. กระตุนการร่วงของใบ ดอก และผล จึงมีชอว่า “Abscisin” ้ ่ื 480 CU 474
69.
6. กระตุนการปิดของปากใบ ้ เอธิ ลีน
(Ethylene) เอธิ ลี น เป็ น แก๊ ซ ที่ พ ื ช สร้ า งขึ้น มาในขณะที่ เ ซลล์ ม ี เ มแทบอลิ ซึ ม ตามปกติ โดยเฉพาะอย่างยิงในขบวนการสุกของผลไม้ นอกจากจะสร้างจากผลแล้วยังมีการสร้างที่ ่ ราก ลําต้น หัว ใบ ดอก และเมล็ด อิ ทธิ พลของเอธิ ลีนต่อการเจริ ญและพัฒนาของพืช 1. กระตุนการหายใจทําให้ผลไม้สกเร็วขึน ผลไม้ทแก่จะสะสมแป้งมาก เอธิลนช่วย ้ ุ ้ ่ี ี เร่งการหายใจทําให้ผลไม้สุก ซึงจะปล่อยแก๊ซเอธิลนออกมาและไปกระตุ้นผลไม้ทอยู่ใกล้ ่ ี ่ี ๆ ให้สกเร็วขึนด้วย ุ ้ 2. ทําให้เกิดการกางของใบ (leaf epinasty) โดยออกซินเป็ นตัวกระตุนให้พชสร้าง ้ ื เอธิลนไปเร่งให้เซลล์ยดตัวทีแตกต่างกัน ี ื ่ 3. ยับยังการพักตัวของเมล็ดและตาในพืชบางชนิด ้ 4. กระตุนการยืดขยายตัวของลําต้น และก้านใบของพืชบางชนิด ้ 5. กระตุนการเกิดรากฝอย (adventitious root) และขนราก (root hair) ้ 6. กระตุนการร่วงของใบและผลโดยกระตุนการแก่ของใบและพัฒนาไปสูการตาย ้ ้ ่ 7. ยับยังการเจริญเติบโตของตาข้าง (Lateral bud) ้ CU 474 481
70.
แบบฝึ กหัดท้ายบท 1. เมื่อพืชดูดแร่ธาตุจากดินเข้าสู่ขนรากแล้วจะถูกลําเลียงผ่านเนื้อเยื่ออะไรบ้าง ก่อนจะถึงวาสคูลาร์บนเดิล ่
ั ิิ ้ 2. พืชใช้ CO2 ในช่วงใดของปฏิกรยาการสังเคราะห์ดวยแสง 3. ออกซิเจนทีเกิดขึนในปฏิกรยาการสังเคราะห์ดวยแสง มาจากโมเลกุลของสาร ่ ้ ิิ ้ ชนิดใด 4. ความหมายทีถูกต้องทีสดของ Dark reaction ในกระบวนการสังเคราะห์ดวย ่ ุ่ ้ แสงเป็ นอย่างไร 5. สารประกอบทีอยูตวชนิดแรกในกระบวนการสังเคราะห์ดวยแสงของพืช คืออะไร ่ ่ ั ้ 6. กลไกการสังเคราะห์ดวยแสง สารทีสร้างขึนในปฏิกรยาใช้แสง (Light reaction) ้ ่ ้ ิิ และถูกนําไปใช้ในปฏิกรยาไม่ใช้แสง คืออะไร ิิ 7. นํ้าตาลตัวแรกทีได้รบจากการสังเคราะห์ดวยแสง คืออะไร ่ ั ้ 8. ผลจากการเกิดปฏิสนธิซอนในพืชดอก ทําให้เกิดสิงใด ้ ่ 9. ผลไม้แบ่งเป็ นกลุมใหญ่ 3 กลุม ได้แก่ อะไรบ้าง ่ ่ 10. ในถุงเอมบริโอ (embryo sac) ของพืช มีนิวเคลียสกีกลุ่ม อะไรบ้าง และนิวเคลียส ่ ทีเกียวข้องกับการปฏิสนธิได้แก่อะไรบ้าง ่ ่ 11. การเคลือนไหวแบบ turgor movement สัมพันธ์กบขบวนการใด ่ ั 12. ถ้าตัดส่วนแผ่นใบออกให้เหลือแต่กานใบ 2-3 วันต่อมา ก้านใบจะหลุดออกจากต้น ้ ทีเป็ นเช่นนี้เกิดจากอะไร ่ 13. การทีรากของพืชพยายามเบนหนีจากแสงสว่างอยูตลอดเวลา เนื่องจากอะไร ่ ่ 14. การทีปลายยอดของพืชเอนเข้าหาแสงสว่างเป็ นเพราะอะไร ่ 15. ฮอร์โ มนพืช ที่ช่ ว ยในการขยายตัว ของเซลล์ลํ า ต้น ช่ ว งระหว่ า งข้อ ถ้ า ขาด ฮอร์โมนนี้จะทําให้ลาต้นเตีย ราบางชนิดสร้างฮอร์โมนนี้ได้ ฮอร์โมนชนิดนี้คออะไร ํ ้ ื 482 CU 474
Download