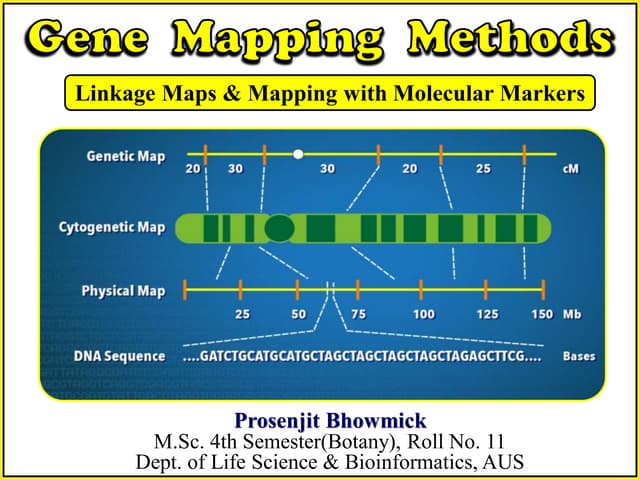
This document discusses bacterial gene mapping techniques. It describes how interrupted conjugation can be used to map genes by determining the order and time at which donor alleles enter recipient bacterial cells. Recombination between donor and recipient DNA during conjugation allows for mapping analysis. Higher resolution mapping can be done by measuring recombinant frequencies between specific genes to determine smaller map distances. Interrupted conjugation experiments provide an initial rough map that is refined through additional experiments measuring recombinant frequencies between different gene combinations.