plastids.pptx
•Download as PPTX, PDF•
0 likes•219 views

plasids

Recommended
More Related Content
What's hot
What's hot (20)
Similar to plastids.pptx
Similar to plastids.pptx (20)
More from Kottakkal farook arts and science college
More from Kottakkal farook arts and science college (20)
Recently uploaded
Recently uploaded (20)
plastids.pptx
- 1. PLASTIDS
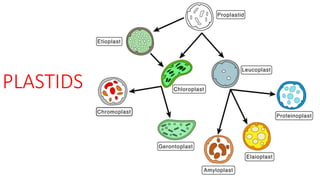
- 2. •Plastids (Schimper - 1883) are self-duplicating membrane-bound cell organelles, concerned with the synthesis and storage of food. •They are present in algae, plants and some protists. •There are three kinds of plastids, namely leucoplasts, chromoplasts and chloroplasts. •They can change from one type to another, and are derived from proplastids. •Leucoplasts are colourless, and the others coloured.Most plastids are coloured due to the presence of pigments. •Some important pigments and their respective colours are given below
- 3. Colour Pigment •1. Chlorophylls Green •2. Carotenes Yellow or red •3. Xanthophylls Yellow, orange or brown •4. Phycoerythrin Red •5. Phycocyanin Blue-green •6. Haematochrome Red •7. Fucoxanthin Brown •8. Bacteriorhodopsin Purple
- 4. •Most pigments are usually seen in plastids. •However, certain other pigments, such as anthocyanins, are found dissolved in the cytoplasmic matrix. •The blue, red, pink and violet colours of some flowers, fruits, stems and leaves are due to the presence of anthocyanins. •Plastids are mainly concerned with the synthesis and storage of food. •Also, they serve asthe centres of several biosynthetic reactions, such as the synthesis of purines, pyrimidines, fatty acids and amino acids.
- 5. LEUCOPLASTS •Leucoplasts (leukoplasts) are colourless plastids, without colouring pigments, thylacoids and ribosomes. •Usually, they are found in germ cells, embryonic cells, meristematic cells and fully differentiated tissues, which are not normally exposed to sunlight. •Plastids, found in cotyledons are initially colourless, but later on they transform to chloroplasts.
- 7. •Leucoplasts are specialised for the synthesis and storage of reserve food. •Leucoplasts which synthesise and store starch are called amyloplasts, those which synthesise and store proteins are called proteoplasts or aleuroplasts, and those which synthesise and store lipids are called elaioplasts (lipidoplasts, oleoplasts, or oleosomes). •Amyloplasts are found in endosperm, cotyledons and storage tubers.
- 8. •Often, leucoplasts get transformed to chloroplasts and chromoplasts under certain circumstances. •The appearance of green colour on stored tubers of potato is due to the conversion of leucoplasts to chloroplasts. •Amyloplasts are found in cotyledons, endosperm and in storage organs, such as potato tubers. •Elaioplasts are commonly found in the tissues of liverworts and monocotyledons. •Aleuroplasts are found in many seeds. •Structurally, leucoplasts are similar to chloroplasts with double layered limiting membrane, but with fewer lamellae.
- 9. CHROMOPLASTS •Plastids, coloured other than green, are called chromoplasts. They occur in the petals, fruits and roots of some higher plants. •Chromoplasts develop either from chloroplasts (as in petals), or rarely from leucoplasts (as in carrot roots). •During the transformation of chloroplast to chromoplast, chlorophylls and starch gradually decrease in quantity, large globuli are formed and arranged along the plastid membrane, lamellar structures break down and stroma gets disorganised. •These changes result in the empty appearance of the plastid centre.
- 11. •During the transformation of leucoplasts to chromoplasts, certain fibrils appear which later on give rise to crystals, filling up the whole plastid. •The crystals are in the form of sheet - like structures, containing large quantities of carotenoids. •Chromoplasts may be red, orange, or yellow. •The red colour of ripening tomatoes is due to the presence of chromoplasts which contain the carotenoid pigment leucopene. •Chromoplasts, containing phycocyanin and phycoerythrin, are found in algae. •In green plants, they contain carotenoid pigments (carotenes and xanthophylls).
- 12. •They show great variety of shape, but are mainly irregular. Granular, angular and forked types also occur. •The irregular and sharply pointed shapes are partly due to the presence of coloured substances in a crystalline form (e.g. roots of carrot). •The chromoplasts of brown algae, dinoflagellates and diatoms, which contain fucoxanthin, are called phaeoplasts. •Those of red algae, which contain phycoerythrin, are called rhodoplasts. •The chromoplasts of blue-green algae (Cyanobacteria) are blue-green in colour and they contain chlorophyll-a, carotenoids and phycobilins.
- 13. CHLOROPLASTS •Chloroplasts are green, chlorophyll-containing, photosynthetically active plastids. •They are the active energy-transducing centres where light energy is trapped, converted to chemical energy and conserved in organic molecules with simultaneous synthesis of ATP. •Chloroplasts are found in green bacteria, green algae, green plants and green protists.
- 15. Morphology •Chloroplasts are among the largest cytoplasmic organelles, clearly observable under the low power of compound microscopes. •Their size, shape and distribution vary with different cells and species. •Their average size varies from 4 to 6 u in diameter and 1 to 3 μ in thickness. •The chloroplasts of polyploid cells may be larger than those of diploid cells. •In general, the chloroplasts of sciophytes (shade-plants) may be larger than those of heliophytes (sun-plants).
- 17. •In some algae, such as Spirogyra, Chlorella and Chlamydomonas, only a single chloroplast is present in each cell. •On the other hand, a cell in the spongy tissue of a grass leaf may have 30 to 50 chloroplasts. •Their average number in land plants is 20 - 40 per cell. •Their division, which occurs in the proplastid stage (immature stage), is not correlated with cell division. •Blue-green algae lack definite chloroplasts; instead they possess loosely arranged cytoplasmic membranes, which contain photosynthetic pigments.
- 18. •Chloroplasts are mostly spherical, ovoid, discoid, vesicular or cup-shaped. •In some algae, they may be reticulate (e.g., Oedogonium), stellate (e.g., Zygnema), band- shaped (e.g., Ulothrix), spiral (e.g., Spirogyra), or cup-shaped (e.g., Volvox). •Chloroplasts are uniformly distributed within the cytoplasm. •Their distribution depends mainly on external conditions, such as light intensity.
- 19. Chemical composition •Chloroplasts are formed of water, proteins, lipids, DNA, RNAs, mineral ions, chlorophylls and other pigments, and the enzymes and biochemical factors required for DNA replication, RNA synthesis, protein synthesis and photosynthesis. •Proteins serve as enzymes and intrinsic and extrinsic membrane proteins. •The lipid contents include phospholipids, triglycerides, sterols and waxes.
- 21. •The photosynthetic pigments are of two groups, namely principal pigments and accessory pigments. •The principal pigment in green plants is chlorophyll-a and those of bacteria include bacteriochlorophyll, chlorobium chlorophyll, bacteriorhodopsin, etc. •The accessory pigments of green plants include chlorophyll-b and carotenoids (carotene and xanthophyll).
- 22. •The accessory pigments of marine algae include chlorophylls-c and d phycoerythrin and phycocyanin. •Accessory pigments expand the light spectrum from which energy can be captured. •The energy absorbed by them is transferred to chlorophyll-a in the early stages of photosynthesis. •Photochemical reaction takes place only in chlorophyll-a.
- 23. Chlorophylls •Chlorophylls are asymmetrical, green, Mg-containing photosynthetic pigments. •They can absorb blue-violet (435-438 nm) and red (670-680 nm) light and transmit or reflect green light (so they appear green). •Their photosynthetic activity will be maximal in red light. •There are different kinds of chlorophylls, such as chlorophylls a, b, c and d, bacteriochlorophyll and chlorobium chlorophyll. •Of these, chlorophylls a and b are found in green plants, and c and d are found in cyanobacteria, some protists and some algae.
- 24. •A chlorophyll molecule is a magnesium-porphyrin derivative. It consists of a pyrole head or nucleus, a phytol tail and a side group. •Head is hydrophilic, and the tail hydrophobic and lipophilic. •Head is a ring of four pyrole molecules, often called tetrapyrole ring or porphyrin ring, with a Mg atom in the centre. •Tail is a long hydrocarbon chain, attached to the porphyrin ring.
- 25. •Linked to the porphyrin ring is a side group, different in different kinds of chlorophylls. •It is the chemical nature of the side group that determines the properties of chlorophyll. •The side group of chlorophyll-a is a methyl group (CH3), and that of chlorophyll-b is an aldehyde group (-CHO). •Chlorophyll a is bluish-green and chlorophyll b is yellowish-green.
- 28. •The empirical formula of chlorophyll-a is C55 H72 O5 N4 Mg, and that of chlorophyll-b is C55 H70 O6 N4 Mg. •Several varieties of chlorophyll - a may be recognized, such as chl.a 673 ,chl.a 683, P680, P700 etc.
- 29. Carotenoids • These are yellow, orange, brown, or red pigments, found in all photosynthetic plants. • Together with anthocyanins, carotenoids impart autumn colours to leaves (when chlorophyll breaks down). • They are found in some flowers and fruits also (e.g., carrot, tomato, pumpkin). • Carotenoids absorb the blue and green light of the visible spectrum and transfer the light energy to chlorophyll to be used in photosynthesis. • They also protect chlorophyll molecules from photo-oxidation (by molecular oxygen in high-intesity light). • Carotenoids are of two groups, namely carotenes (C40H56) and xanthophylls (C40H56O₂).
- 30. • Carotenes are red or orange-coloured hydrocarbons (terpenes). • Xanthophylls are brown or yellow and oxygenated hydrocarbons. • The common carotenes include a, ß, y and d carotenes, phytotene, lycopene, neurosporene, etc. • During digestion in vertebrates, B-carotene gets hydrolysed to two identical portions, which yield vitamin A. • Xanthophylls are more abundant than carotenes. • They differ from carotenes in having oxygen. • The major xanthophylls of higher plants include lutein, violaxanthin, zeaxanthin and neoxanthin. • The commonest xanthophyll of diatoms and brown algae is fucoxanthin. • Lutein gives yellow colour to autumn leaves.
- 31. Phycobilins • Phycobilins are red or blue accessory photosynthetic pigments, found in cyanobacteria and red algae. • They differ from chlorophylls and carotenoids in being water-soluble. • They are similar to the porphyrin part of chlorophylls, except that Mg is absent and the tetrapyrroles are linear rather than cyclic. • There are two major groups of phycobilins, namely phycoerythrins and phycocyanins. • Phycoerythrins are red-coloured. • They absorb dim and blue-green light, which can reach ocean depths. • So, phycobilins enable algae to live in deep waters. • Phycocyanins are bluecoloured. • They absorb extra orange and red light.
- 32. Bacteriorhodopsin • In Halobacteria, there is a light-absorbing proteinous pigment in the plasma membrane, called bacteriorhodopsin. • It is formed of a single polypeptide of 247 amino acids. • It resembles the visual pigment rhodopsin, found in the retinal rods of vertebrate eye. • It exists in two interconvertible forms, namely "purple" and "bleached". • The former absorbs light at a wavelength of 570 nm, and the latter at 412 nm. • As the pigment transforms from the purple to the bleached form, it loses H+ ion, and in the reverse transformation it picks up and H+ ion (Racker and Stoeckenius 1974). • Thus, bacteriorhodopsin functions as a proton pump, that is driven by light.
- 33. Structure of chloroplast • Chloroplast is a double-walled and fluid-filled bag. • It has a gelatinous core, called matrix or stroma, covered by two concentric membranes. • Across these membranes, molecular exchange takes place between stroma and cytosol. • The outer membrane is highly permeable, and the inner one is less permeable. • In between these membrane is the intermembrane space or periplastidial space. • The membranes have lipoprotein composition, lamellar structure and fluid-mosaic organization. • They consist of two separate layers of 40-60 Å thickness.
- 35. • Stroma is a metabolic centre where CO₂ fixation and the synthesis of nucleic acids, sugars, starch, fatty acids and some chloroplast proteins take place. • It contains a lamellar membrane system, ribosomes, circular DNA, messenger and transfer RNAs, starch grains, lipid globules (called plastoglobuli), and the enzymes for DNA duplication, genetic transcription, genetic translation and the dark reactions of photosynthesis. • Stroma contains nearly 50% of the chloroplast proteins most of which are of the soluble type. • Most algal chloroplasts contain pyrenoids, in association with starch grains. • Pyrenoids contain the active enzyme ribulose biphosphate carboxylase. • Hence, they are believed to be involved in carbohydrate synthesis.
- 36. •In lower organisms, chloroplasts are primitive. •They are bounded by a two-layered membrane and contain pigment-coated plates in the matrix. Such chloroplasts are called lamellate chloroplasts. •The extensive stromal membrane system is called thylacoid system or lamellar system. •Chlorophyll molecules and light-absorbing accessory pigments are embedded in it. It mainly consists of several cylindrical discs, called grana. •Each granum is a parallel stack or pile of 5 to 50 or more flattened, closed and pigmented sacs, called granal thylacoids or granal lamellae.
- 39. •Adjacent grana are connected together by a system of tubular membranes, called intergranal thylacoids or fret membranes. •In addition to these, a few unstacked thylacoids may also be present in the stroma. •They are called stromal thylacoids or stromal lamellae. •They form a system of anastomosing tubules that are joined to the grana thylacoids •The number of thylacoids in a granum is different in different groups. •There is only a single thylacoid in each granum in Rhodophyceae, two in Cryptophyceae, three in Phaeophyceae and Bacillariophyceae, and many in others.
- 40. •In the bundle sheath cells of C4 plants, a specialised type of chloroplasts are found. •In them, lamellae are uniformly dispersed in the stroma, without forming grana. So, they are called agranal chloroplasts. •The leaves of C4 plants have the peculiar kranz anatomy and hence their agranal chloroplasts are also called kranz type of chloroplasts. •Different views have been expressed to explain the relation between granal and intergranal thylacoids. •Steinmann and Sjostrand hold that stromal and intergranal lamellae develop from granal discs. Hedge and others are of opinion that granal discs are locally swollen thylacoids.
- 41. •Thylacoids have lipoprotein composition and laminar organization. •They are formed of thin molecular sheets or layers, called laminae or lamellae. •These are the store houses of photosynthetic pigments and the enzymes of the light reactions of photosynthesis. •The membranes also contain photochemically inactive light-harvesting complex proteins (LHCP) and the electron carriers cytochrome-b, cytochrome-f, plastoquinone and plastocyanin.
- 42. Photosystems or pigment systems •The light-absorbing photosynthetic pigments in the thylacoid membrane are arranged in two complex pigment systems, namely photosystem I(PSI) and photosystem II (PSII). •PSI is present mainly in unstacked membranes, and PSII mainly in stacked membranes. •These two systems are structurally distinct, but functionally related. They are linked together by mobile electron carriers.
- 43. •Each photosystem has many thousand functional photosynthetic units, called quantasomes. •Each unit, in turn, consists of nearly 200-400 chlorophyll molecules and many accessory pigment molecules. •The pigment systems can absorb all wavelengths of the visible spectrum, especially those between 400 and 500 nm, and those between 600 and 700 nm.
- 44. •Each functional unit has a photochemical reaction centre, often called photocentre. •It serves as an energy trap for the collection and conversion of light energy; it brings about the conversion of light energy to chemical energy. •It is formed of chlorophyll-a molecules, bound to a protein complex. •The reaction centre of PSI is termed P700 and that of PSII is termed P680. •The former has an absorption maximum at 700 nm, and the latter has it at 680 nm.
- 45. •The reaction centre is surrounded by a cluster of light- absorbing pigment molecules. •They form a light-gathering antenna system to absorb photons. •Hence, they are often called light-harvesting antenna molecules. •They include chlorophyll-a, chlorophyll-b, ß-carotenes and other accessory pigments.
- 46. • Antenna molecules remain complexed with a few polypeptides, forming the light-harvesting complex (LHC). • This complex is more prominent in PSII than in PSI. • It is mainly localized in stacked thylacoids. Its main function is to capture solar energy and it has no photochemical activity at all. • PSI and PSII differ from each other with regard to their LHC; in the LHC of PSII, the amount of chlorophyll-a is much lesser than those of chlorophyll-b and ß-carotene, while in the LHC of PSI it is much higher than those of chlorophyll-b and ßcarotene. • In the photosystems, the pigment molecules are very closely and orderly arranged. • This enables each molecule to absorb photon energy and pass it on to the next molecule and finally to the reaction centre by a chain- transfer mechanism.
- 47. Quantasomes • Quantasomes are closely packed arrays of spherical or oblate granular particles, regularly arranged on or in chloroplast thylacoids. • They are considered to be the smallest light harvesting photosynthetic units, capable of carrying out photochemical reactions. • In green plants, each of them contains nearly 200-400 chlorophyll-a molecules, some accessory pigments, electron carriers and the enzymes of light reactions. • [Quantasomes were first noticed by Moor, Weier and Benson, and were first isolated by Park and Biggins-1964] • Evidences suggest that at least two distinct classes of quantasomes are present, namely the smaller ones on the membrane surface and the larger ones inside the membrane. • Functionally, the smaller and larger quantasomes are believed to represent pigment systems (photosystems) I and II respectively.
- 48. Semigenetic autonomy of chloroplasts •Cytological studies have revealed the presence of circular DNA, RNAs and ribosomes in chloroplasts. •Nucleic acids are of the bacterial type. •The DNA content of a chloroplast is about ten times greater than that of a mitochondrion. •The presence of DNA, RNAs and ribosomes gives chloroplasts the ability to synthesise proteins. •This makes them genetically semiautonomous.
- 49. Functions of chloroplasts •(i) Energy fixation and the synthesis of organic molecules: • Chloroplasts are the centres of photosynthesis. • By photosynthesis, they manufacture organic molecules and bring about energy fixation. • Energy fixation is the overall process by which solar energy is trapped, converted to chemical energy and conserved in the energy-bonds of organic molecules. • Energy fixation is significant in that it brings about the transduction of light energy to chemical energy and thereby initiates the dynamics of the ecosystem. • Thus, the functional state of the ecosystem seems to be largely dependent on the energy fixation by chloroplasts.
- 50. (ii) Synthesis of nucleic acids and proteins: • Just as mitochondria, chloroplasts are also semiautomous organelles. • With some autonomy they too can synthesise nucleic acids and protein and thus bring about gene expression through the replication, transcription and translation of the genetic code. • Chloroplasts contain the complete molecular machinery for the synthesis of nucleic acids and proteins. • Chloroplast DNA can underego enzymatic replication and produce multiple copies of it. • Similarly, it can undergo transcription and synthesise RNAs. • Making use of their DNA, RNAs, ribosomes and enzymes chloroplasts can carry out genetic translation and synthesise some chloroplast proteins also.
- 51. (iii) Extra-chromosomal (cytoplasmic) inheritance: •Chloroplast DNA contains extra-chromosomal genes, called plastogenes or plasma genes. •They can govern non-Mendelian inheritance and control plastid characters, generally in co- operation with chromosomal genes. •Plastogenes are believed to control the inheritance of leaf colour in some plants.
- 52. (iv) Provide energy and carbon for all heterotrophs: •Chloroplasts carry out photosynthesis and produce organic molecules, which form the basic source of energy and carbon for all heterotrophic organisms. •So, most organisms directly or indirectly depend on chloroplasts for their energetic and nutritional requirements.
- 53. v) Enrich atmospheric O, and maintain the stratospheric ozone layer: •Oxygenic photosynthesis in chloroplasts removes CO₂ from the atmosphere and releases O₂ to the atmosphere. •This enriches the atmospheric oxygen store. •Virtually, the entire atmospheric oxygen build up is the product of oxygenic photosynthesis over the past 3300 million years. •A portion of this oxygen treasure is converted to stratospheric O, by the action of UV radiation.
- 54. •This ozone layer screens out harmful solar and cosmic radiations and thereby protects the life on earth. •Thus, it becomes clear that without chloroplasts and green plants there would be no oxygen in the atmosphere, and life on earth would be almost impossible.
- 55. (vi) Reduction of nitrite to ammonia : •The reducing power of energised (light- activated) electrons in chloroplasts drives the reduction of NO₂ to NH3. •The ammonia thus formed provides nitrogen for the synthesis of nitrogenous compounds, such as amino acids, nucleotides, etc.
- 56. Origin of plastids • Many theories have been put forward to explain the origin and development of plastids. Some of them are the following: (a) Monotropic development • This theory holds that plastids are autonomous organelles and they can change from one form to another either reversibly or irreversibly. • So, according to it, plastids are not formed de novo, but are always formed by interconversion. • It also holds that chromoplast is a degenerated form of chloroplast, formed by the irreversible transformation of chloroplast. • This transformation involves an irrecoverable loss of chlorophyll.
- 57. b) Development from proplastids • Several workers hold that plastids develop from tiny cytoplasmic bodies, called proplastids or plastid precursors. • Proplastids are simple self-duplicating structures with a clear matrix, bounded by a double-layered membrane. • The matrix contains DNA and RNAs.Proplastids develop first in the cytoplasm of meristematic cells. • On exposure to light, they elongate and develop 'blebs' from the inner side of their double-layered envelope. • These blebs become lamellae, which in turn, multiply through linear splitting. • Simple lamellae undergo growth and differentiation and give rise to granal, intergranal and stromal lamellae. This results in the transformation of proplastids to mature chloroplasts.
- 58. (c) Division and budding •In lower plants, new plastids originate by the division and budding of the existing ones. •During division, ring-like constrictions appear, which gradually deepen and finally divide the plastid into fragments. •These fragments then differentiate into mature plastids. •Budding of plastids also takes place under certain special circumstances.
- 59. (d) Nuclear origin •Proplastid-like structures have been observed near the nucleus in some cases. •This has led some workers to think that proplastids originate from nucleus by the evagination of nuclear envelope. •Then, proplastids develop to mature plastids.