golgi bodies.pptx
•Download as PPTX, PDF•
0 likes•51 views

golgi bodies

Recommended
More Related Content
Similar to golgi bodies.pptx
Similar to golgi bodies.pptx (20)
More from Kottakkal farook arts and science college
More from Kottakkal farook arts and science college (20)
Recently uploaded
Recently uploaded (20)
golgi bodies.pptx
- 1. GOLGI BODIES
- 2. •Golgi bodies are aggregations of membrane-bound spaces, which serve as the centres of cellular secretion. •They are particularly abundant in secretory cells. •They form a specialised part of the endomembrane system, and serve as the centres of cellular secretion. •They are primarily concerned with the concentration, chemical modification and packaging of synthetic products as secretory granules or droplets.
- 3. DISCOVERIES • Early observations of Golgi bodies were made by St. George, Plattner, Hermann and others. • But, Vacuoles they were first discovered and described by Camillo Golgi (1898). • He termed them "apparato reticulare interno" (internal reticular apparatus). • Since then, they came to be known variously as Golgi bodies, G. complex, G. apparatus, G. vesicles, and so on. • Baker (1951) proposed the name lipochondrins, considering their presumed lipid content. • Plant Golgi bodies are often described as dictyosomes.
- 5. • Golgi bodies occur in all kinds of cells, excepting prokaryotes and mature mammalian RBCs. • Their location, number and size are different in different cell types. • In secretory cells, they are usually seen in between the nucleus and the apical pole. • But in nerve cells, they occupy a circum-nuclear position. • In most cells, there is only a single set of Golgi bodies. • But, invertebrate cells, nerve cells, liver cells and most plant cells have multiple sets, more or less distributed throughout the cytoplasm. • Multiple sets of Golgi bodies are described dictyosomes. • In salivary gland cells, they are exceptionally numerous, often several thousand.
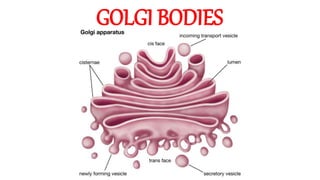
- 8. Morphology • Golgi bodies are essentially membrane-bound spaces. • They are of three kinds, namely lamellae, vesicles and vacuoles. • Lamellae form the core units of Golgi bodies. • Vesicles and vacuoles remain crowded around them in clusters. • The internal lumen of lamellae is called cisterna. • Lamellae are flattened sacs, with dilated ends. • They are arranged in parallel stacks or piles, often called golgiosomes.
- 10. •Vesicles are groups of small membrane-bound spaces, and vacuoles are large and scattered membrane-bound sacs. •Vesicles, vacuoles and the dilated ends of lamellae may be filled with a granular or amorphous material. •In some cases, the spaces between adjacent lamellae (interlamellar spaces) may also be filled with a similar material. •Its composition and functions are not definitely known. •Probably, it may serve as a cementing substance.
- 11. •Lamellar membranes are always in dynamic equilibrium. •They are constantly budded off from the SER and, at the same time, lost for the formation of secretory vesicles. •In goblet cells, they get transformed to mucilage granules, which almost fill the whole cell. •Each lamella has a polarised orientation. •It has a convex outer side or face, known as the "forming face", entry face, or cis face, and a concave inner face, known as "maturing face" (exit face or trans face).
- 12. •The forming face receives vesicles from ER, whereas maturing face gives out a large number of secretory vesicles. •The forming face is oriented towards RER or nuclear envelope, and the maturing face towards the cell surface. •Each face is closely associated with a network of tubules, forming the so called cis Golgi network and trans Golgi network.
- 13. •A lamellar stack contains three major kinds of sacs with different functions. They are cis sacs, trans sacs and medial sacs. •Cis sacs are located towards the outer or cis face of the stack, trans sacs are located towards the inner or trans face, and medial sacs are seen in the middle of the stack. •Cis sacs are mainly concerned with the phosphorylation of the mannose molecules of the glycoproteins that are destined to pass to lysosomes. •This phosphorylation serves as a specific signal to direct proteins to lysosomes.
- 14. •Medial sacs are involved in the removal of mannose molecules and the addition of acetyl glucosamine to the glycoproteins that are destined for exocytosis through plasma membrane. •The removal of mannose molecules is mediated by the enzymes mannosidases, and the addition of acetyl glucosamine is mediated by glycosyl transferases. •Trans sacs are involved in the addition of galactose and sialic acid (a modified hexose sugar) to glycoproteins. •The transport of glycoproteins from cis-to-medial-to-trans sacs is effected by the vesicles which bud from one sac and then fuse with the next sac.
- 15. • In between the forming face and the RER, there are numerous membrane-bound bodies, called transition vesicles or transition tubules. • Often, they converge upon the lamellae and form a fenestrated plate. • These transition elements are thought to take their origin from RER and, by fusion, they form new lamellae. • This compensates the loss of lamellae at the maturing face. • Associated with the maturing face, there may be present a saccular region of SER, known as GERL.
- 16. • The vesicles of the Golgi complex occur in two forms, namely smooth-surfaced vesicles and rough-surfaced or "coated vesicles". • Smooth vesicles are quite common, but rough vesicles are less frequent. • The membranes of the rough vesicles are coated with granules or bristles at the outer (cytoplasmic) surface. • The functional significance of this morphological distinction is not definitely understood. • Golgi bodies have no attached ribosomes. • Their membranes appear to be surrounded by a small area, often called the zone of exclusion. • Here, glycogen, ribosomes and mitochondria are apparently absent.
- 17. Chemical composition •Golgi bodies are chemically unstable. •Their chemical make-up may change from time to time. •In general, their membranes have lipoprotein composition, laminar structure and fluidmosaic organization. •They are formed mainly of proteins and phospholipids, most usually in the ratio 6:4, or sometimes in almost equal amounts. •The presence of low levels of cholesterol, cholesterol esters, carotenoids, free fatty acids, triglycerides, mucopolysaccharides, vitamin C, etc. has also been reported in several cases.
- 18. •Golgi bodies are rich in a variety of enzymes. •The enzymes, found in high concentrations, include thiamine pyrophosphatases and glycosyl transferases. •Transferases are the most characteristic Golgi enzymes, and they mediate the synthesis of glycoproteins and glycolipids by transferring oligosaccharides to proteins and lipids. •Some acid phosphatases (such as ADPase, ATPase and CDPase), phospholipases, oxido-reductases, transferases, and some lysosomal enzymes may also be found in Golgi bodies.
- 20. Biogenesis • The components of the Golgi complex are constantly re-formed in a more or less dynamic and cyclic manner • Different views have been advanced to account for the origin of Golgi membranes. • One view is that they arise de novo (anew). • The second view is that they arise from pre-existing membranes, mainly from plasma membrane, ER and nuclear membrane. • The third view is that Golgi lamellae take their origin from SER and then they bud off the other components. • The SER is believed to take its origin from RER which, in turn, is formed from nuclear membrane.
- 21. Functions •Golgi bodies are the important chemical factories or secretory centres of the cell. •They are involved in the synthesis of substances, storage, processing and packaging of secretory products as secretory vesicles, formation of acrosome and primary lysosome, activation of mitochondria, elaboration of yolk bodies, etc. •Some of their major functions are the following: •(i) Serve as an intracellular compartment for the chemical modification of the substances synthésised elsewhere, and also for the differentiation and rapid turn over of cellular membranes.
- 22. • (ii) Synthesis of mucopolysaccharides, glycoproteins and glycolipids by glycosylation (addition of sugar residues from a sugar donor). This takes place in a step by step manner with the help of the enzymes glycosyl transferases. The donor of sugar units in most cases is uridine diphosphate glucose (UDPG). • (iii) Cellular secretion of export proteins, intracellular enzymes (such as lysosomal and peroxisomal enzymes), hormones, antibodies, yolk granules, lipid droplets, etc. • (iv) Secretion of mucus, milk protein, thyroxine compounds, etc. • (v) Formation and packaging of zymogen granules, melanin granules, etc. • (vi) Formation of microbodies and primary lysosomes and the glycosylation of lysosomal enzymes.
- 23. • (vii) Formation of acrosome in developing animal sperms, and the formation of cortical granules and a protective coating in developing oocytes. • (viii) Removal and re-cycling of excess membranes and secretory granules (e.g., zymogen granules). •(ix) Formation, packaging and transport of cell wall polysaccharides and pectic compounds for cell wall formation in plant cells. •(x) Formation of cell plate during the division of plants cells. This is accomplished by the condensation of membrane vesicles which discharge their contents to reinforce the cell plate.