![Maintaining Homeostasis ,[object Object],[object Object],[object Object],[object Object],[object Object],[object Object]](data:image/gif;base64,R0lGODlhAQABAIAAAAAAAP///yH5BAEAAAAALAAAAAABAAEAAAIBRAA7)
Recommended
More Related Content
What's hot
What's hot (20)
Viewers also liked
Viewers also liked (20)
Similar to Membranes pt. 2
Similar to Membranes pt. 2 (20)
More from Jolie Yu
More from Jolie Yu (20)
Recently uploaded
Recently uploaded (20)
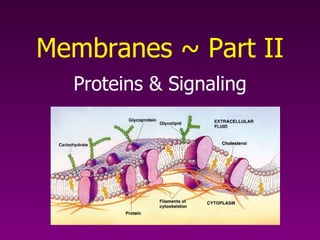
Membranes pt. 2
- 1. Proteins & Signaling Membranes ~ Part II
- 4. Mechanisms of Cell Communication
- 9. Ligand Gated Ion Channel
- 18. An Overview of Cell Signaling
- 26. Tyrosine Kinase Receptor Dimers
- 30. Exchange of Yeast Mating Factors
- 43. Cyclic AMP
- 45. cAMP Second Messenger System