1. The document discusses the metabolism of purine and pyrimidine nucleotides, including their biosynthesis, degradation, and disorders related to purine metabolism such as gout and Lesch-Nyhan syndrome.
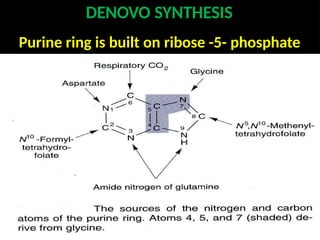
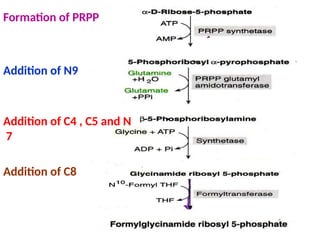
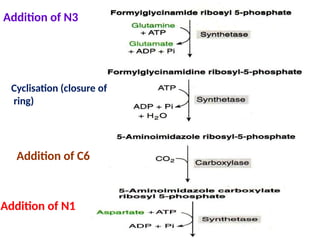
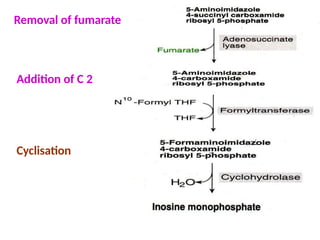
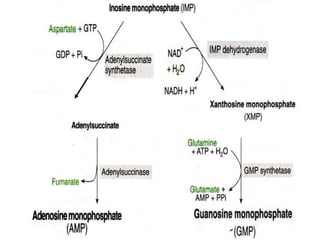
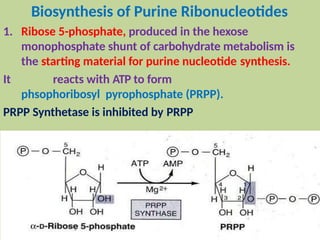
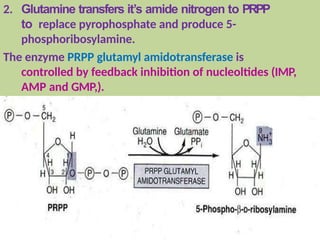
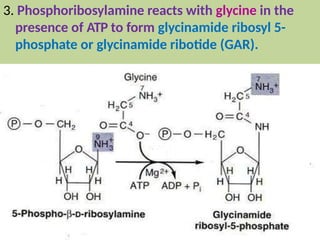
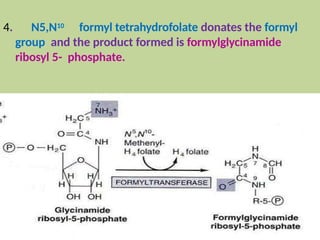
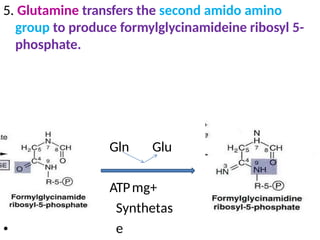
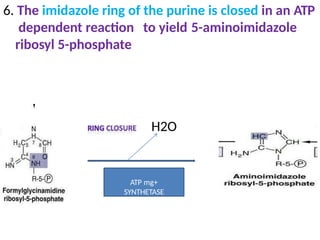
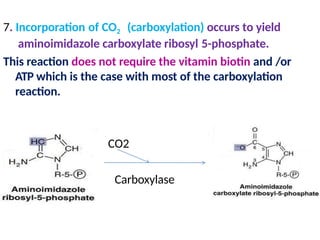
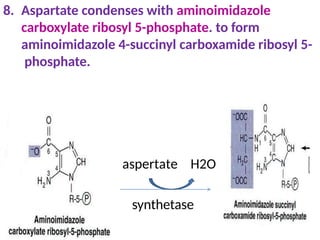
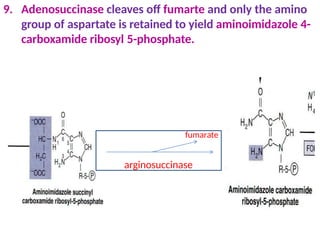
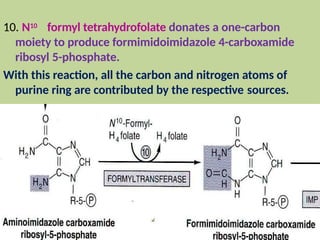
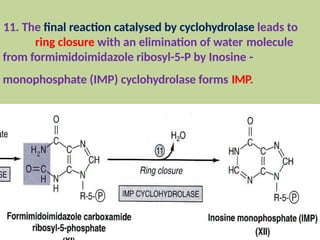
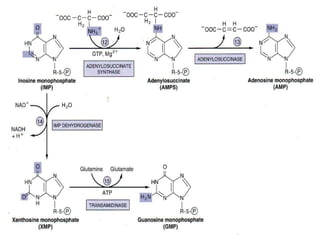
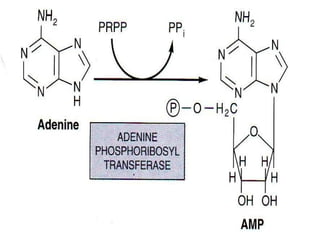
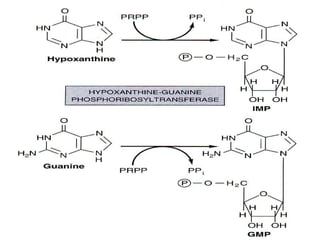
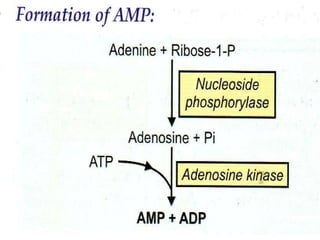
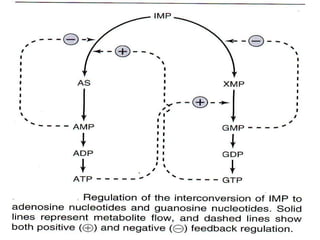
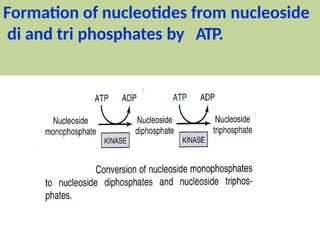
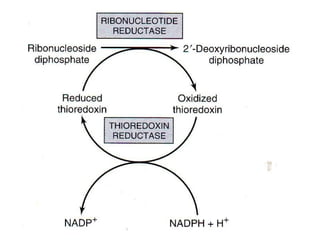
2. Purine nucleotides are synthesized through both a de novo synthesis pathway that builds the purine ring from simple precursors, and a salvage pathway that recycles purine bases. A key early intermediate is IMP, which is converted to AMP and GMP.
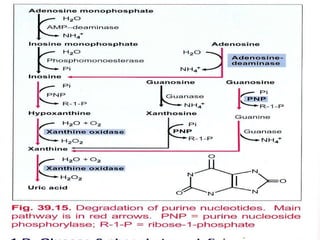
3. Uric acid is the final product of purine degradation in humans and high levels can cause gout. Lesch-Nyhan syndrome results from HGPRT deficiency disrupting the salvage pathway.























































![PEPTIDOMIMETICS [M.PHARM, BSC, MSC, B.PHARM]](https://cdn.slidesharecdn.com/ss_thumbnails/53-191219102328-thumbnail.jpg?width=640&height=640&fit=bounds)
















































