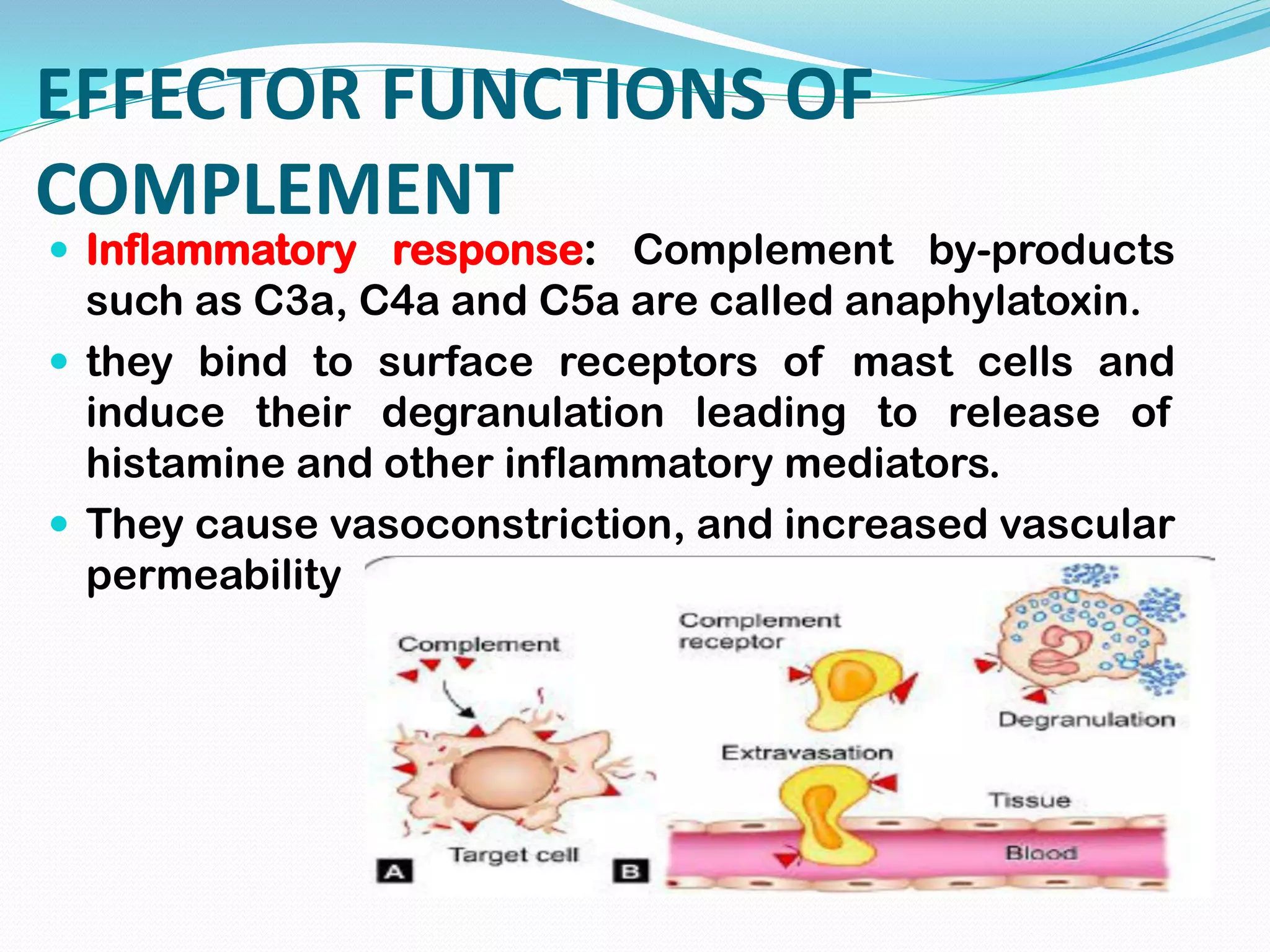
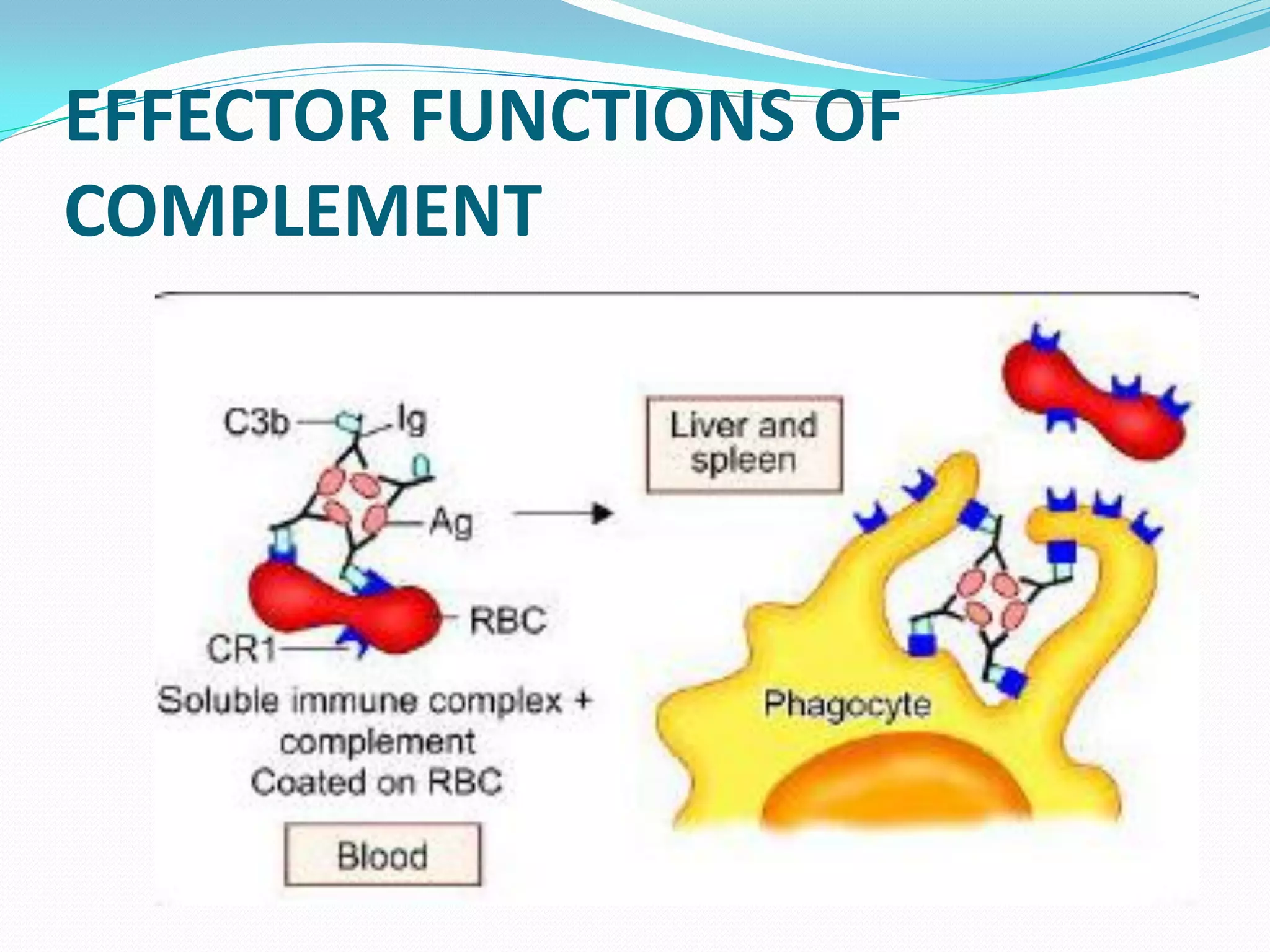
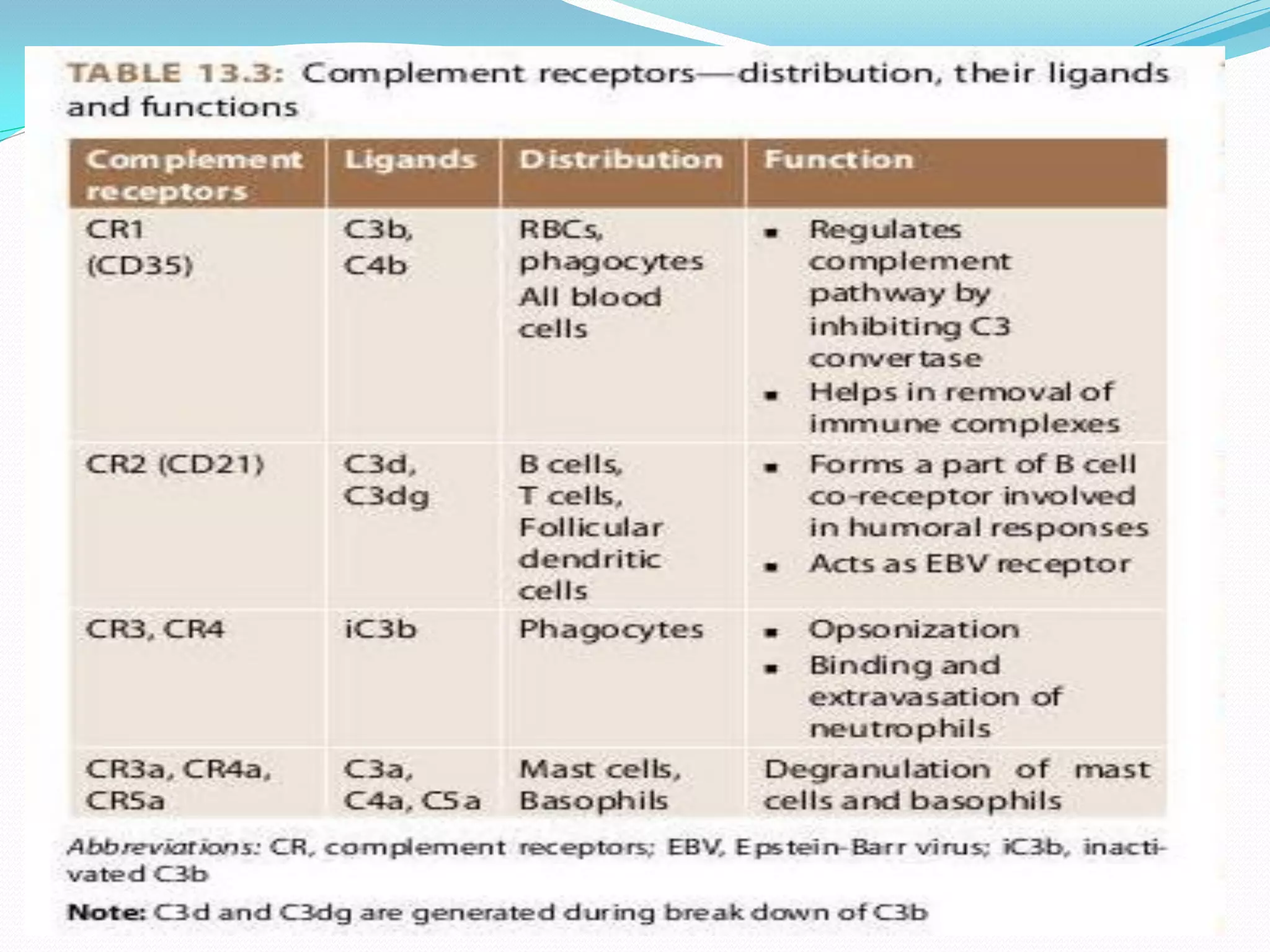
The complement system comprises over 30 proteins that augment the immune response. It has three pathways - classical, lectin, and alternative. The classical pathway is antibody-dependent and initiates with C1 binding to antigen-antibody complexes. The lectin pathway involves mannose-binding lectin and is antibody-independent. The alternative pathway is also antibody-independent and initiates with C3 binding directly to pathogens. All three pathways form C3 and C5 convertases and the membrane attack complex (MAC) to lyse target cells. Complement effectors also mediate inflammation and opsonization. The system is tightly regulated to prevent damage to host cells. Deficiencies can increase susceptibility to infection.