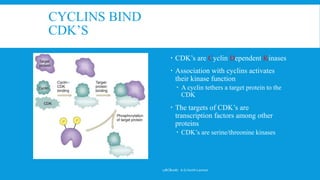
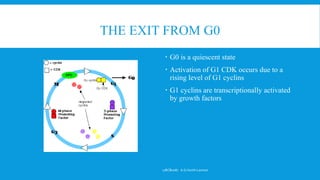
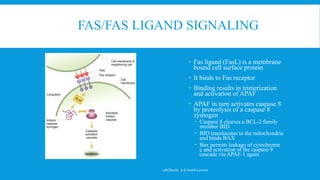
The document discusses the processes of cell cycle and apoptosis, detailing how cells transition through division, quiescence, and programmed death. It emphasizes the roles of cyclins and cyclin-dependent kinases (cdks) in regulating the cell cycle, checkpoints, and mechanisms of apoptosis, distinguishing it from necrosis. Key players in these processes include p53, which regulates DNA damage responses and can induce apoptosis, along with various signaling pathways that lead to cell death.








































![Cell cycle. sgd[1]](https://cdn.slidesharecdn.com/ss_thumbnails/cellcycle-150228084926-conversion-gate01-thumbnail.jpg?width=640&height=640&fit=bounds)












































