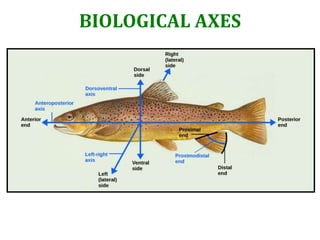
Biological Axes of Regeneration
•Download as PPTX, PDF•
1 like•980 views

This document discusses regeneration in various organisms including salamanders, hydra, and planaria. It describes two main types of regeneration - epimorphosis and morphallaxis. Epimorphosis involves cellular proliferation, dedifferentiation, and blastema formation while morphallaxis involves tissue remodeling without proliferation. Salamanders undergo epimorphic regeneration of limbs through formation of a regeneration blastema. Hydra regenerate through morphallaxis and some epimorphosis, regulated by Wnt signaling and head/foot activation gradients that determine where new structures form.

Recommended
More Related Content
What's hot
What's hot (20)
Similar to Biological Axes of Regeneration
Similar to Biological Axes of Regeneration (20)
More from SABARI KRISHNAN B. B.
More from SABARI KRISHNAN B. B. (6)
Recently uploaded
Recently uploaded (20)
Biological Axes of Regeneration
- 3. Regeneration is the re-activation of development in post-embryonic life to restore missing or damaged tissues of an organism. Although regeneration takes place in nearly all species, several organisms have emerged as particularly fruitful models for the study of regeneration. Hydra and Planaria are able to regenerate into complete organisms following amputation or from very small fragments. Certain salamanders are unique among tetrapods in being able to regenerate whole limbs, and frog larvae are often used to study the regeneration of the tail and the lens of the eye. Zebrafish have recently proved advantageous for investigating the mechanisms of regeneration of CNS, retina, heart, liver and fin. Although mammals are unable to rebuild whole appendages, individual tissues and organs do possess variable regenerative capabilities; most notable are the antlers of deer.
- 4. Thomas Hunt Morgan named the two parts of the regenerative process epimorphosis and morphallaxis. Specifically, Morgan wanted epimorphosis to specify the process of entirely new tissues being regrown from an amputation or similar injury, with morphallaxis being coined to describe regeneration that did not use cell proliferation, such as in hydra.The key difference between the two forms of regeneration is that epimorphosis involves cellular proliferation and blastema formation, whereas morphallaxis does not. Two main classifications: 1. Epimorphosis. In some species, adult structures can undergo dedifferentiation to form a relatively undifferentiated mass of cells (a blastema) that then re-differentiates to form the new structure. E.g. regenerating amphibian limbs. 2. Morphallaxis. Here, regeneration occurs through the re-patterning of existing tissues (transdifferentiation), and there is little new growth. E.g. regeneration in Hydra. Other classifications: Stem cell-mediated regeneration. Stem cells allow an organism to regrow certain organs or tissues that have been lost. E.g. regrowth of hair shafts from Follicular SCs in the hair bulge and the continual replacement of blood cells from HSCs. Compensatory regeneration. Here the differentiated cells divide but maintain their differentiated functions. The new cells do not come from stem cells, nor do they come from the dedifferentiation of the adult cells. Each cell produces cells similar to itself; no mass of undifferentiated tissue forms. E.g. regeneration of mammalian liver.
- 6. Epimorphosis : the regeneration of a specific part of an organism in a way that involves extensive cell proliferation of somatic stem cells, dedifferentiation, and reformation, as well as blastema formation. Epimorphosis can be considered a simple model for development, though it only occurs in tissues surrounding the site of injury rather than occurring system-wide. Epimorphosis restores the anatomy of the organism and the original polarity that existed before the destruction of the tissue and/or a structure of the organism. Epimorphosis regeneration can be observed in both vertebrates and invertebrates such as salamanders, annelids and planarians. SALAMANDERS: EPIMORPHIC LIMB REGENERATION • When an adult salamander limb is amputated, the remaining limb cells reconstruct a new limb, complete with all its differentiated cells arranged in the proper order. Remarkably, the limb regenerates only the missing structures and no more. For example, when the limb is amputated at the wrist, the salamander forms a new wrist and foot, but not a new elbow. In some way, the salamander limb “knows” where the proximal-distal axis has been severed and is able to regenerate from that point on. • Salamanders accomplish epimorphic regeneration by cell dedifferentiation to form a regeneration blastema, which in this case is an aggregation of relatively undifferentiated cells derived from the originally differentiated tissue that then proliferates and re-differentiates into the new limb parts. • Bone, dermis, and cartilage just beneath the site of amputation contribute to the regeneration blastema, as do satellite cells from nearby muscles. Hence much of the salamander limb’s regeneration blastema appears to arise from the dedifferentiation of adult cells followed by cell division and the re-differentiation of those cells back into their original cell types.
- 8. Formation of the apical epidermal cap and regeneration blastema • When a salamander limb is amputated, a plasma clot forms. • Within 6–12 hours, epidermal cells from the remaining stump migrate to cover the wound surface, forming the wound epidermis. In contrast to wound healing in mammals, no scar forms, and the dermis does not move with the epidermis to cover the site of amputation. • The nerves innervating the limb degenerate for a short distance proximal to the plane of amputation. • During the next four days, the extracellular matrices of the tissues beneath the wound epidermis are degraded by proteases, liberating single cells that undergo dramatic dedifferentiation: bone cells, cartilage cells, fibroblasts, and myocytes all lose their differentiated characteristics. • Genes that are expressed in differentiated tissues (such as the mrf4 and myf5 genes expressed in muscle cells) are down-regulated, while there is a dramatic increase in the expression of genes such as msx1 that are associated with the proliferating progress zone mesenchyme of the embryonic limb. • This cell mass is the regeneration blastema, and these cells are the ones that will continue to proliferate and that will eventually re-differentiate to form the new structures of the limb. • Moreover, during this time, the wound epidermis thickens to form the apical epidermal cap (AEC), which acts similarly to the apical ectodermal ridge during normal limb development. • Kragl and colleagues (2009) found that the blastema is not a collection of homogeneous, fully dedifferentiated cells. Rather, in the regenerating limbs of the axolotl salamander, muscle cells arise only from old muscle cells, dermal cells come only from old dermal cells, and cartilage can arise only from old cartilage or old dermal cells. Thus, the blastema is not a collection of unspecified multipotent progenitor cells. Rather, the cells retain their specification, and the blastema is a heterogeneous assortment of restricted progenitor cells.
- 10. Proliferation of blastema cells • The growth of the regeneration blastema depends on the presence of both the AEC and nerves. • The AEC stimulates the growth of the blastema by secreting Fgf8, but the effect of the AEC is only possible if nerves are present. • Both sensory and motor axons in a threshold amount should innervate the blastema such that sensory axons make direct contact with the AEC and motor axons terminate in the blastema mesenchyme. • Most important is that regenerating nerves are necessary for the proliferation and outgrowth of the blastema. • If the limb is first enervated and then amputated, no regeneration will occur. • If a wound is made in the epidermis of the proximal limb and a nerve is then diverted to the wound area, a blastema-like bud will form, but not a fully regenerated limb. To induce a complete ectopic limb, not only does a nerve need to be diverted to the wound site, but an epidermal graft from the opposite side of the limb (from a posterior to an anterior location) needs to be placed near the wound. These results suggest that during normal limb regeneration, the regenerating nerves deliver important signals to the AEC. They also suggest, however, that signals from nerves are not sufficient for ectopic limb growth; for that growth, positional cues from an epidermis that are different from the positional cues at the wound site itself are also needed.
- 11. • However, experiments have shown that neural conductance (action potentials and release of acetylcholine) is not required to promote limb regeneration. If neural activity is not required, then what are the regenerating axons providing to the limb blastema? These neurons are believed to release factors necessary for the proliferation of the blastema cells. • There have been many candidates for such a nerve-derived blastema mitogen, but probably the best candidate is newt anterior gradient protein (naG). This protein can cause blastema cells to proliferate in culture, and it permits normal regeneration in limbs that have been enervated. • If activated nAG genes are electroporated into the dedifferentiating tissues of amputated limbs that have been enervated, the limbs are able to regenerate. If nAG is not administered, the limbs remain stumps. Moreover, nAG is only minimally expressed in normal limbs, but it is induced in the Schwann cells that surround the regenerating axons within 5 days of amputation. • When researchers compared the expression of nAG of normal limbs with that of both enervated limbs and these aneurogenic limbs, they discovered that the aneurogenic limbs had a uniquely high level of nAG expression in the epidermis. Moreover, upon limb amputation, nAG increased first in the nerve sheath of normal limbs, but was present throughout the blastema in the aneurogenic limbs. These results suggest that nAG alone is the primary mitogen responsible for nerve-dependent regeneration. The receptor for nAG, Prod1, has since been discovered and found to be expressed in a proximal-to-distal gradient in the salamander limb. This ligand- receptor relationship seems to be conserved across regeneration-competent salamander species.
- 14. • Morphallaxis : the regeneration of specific tissue in a variety of organisms due to loss or death of the existing tissue. The classical example of morphallaxis is that of the cnidarian Hydra, where when the animal is severed in two, the remaining severed sections form two fully functional and independent hydra. • The notable feature of morphallaxis is that a large majority of regenerated tissue comes from already- present tissue in the organism. That is, the one severed section of the hydra forms into a smaller version of the original hydra, approximately the same size as the severed section. Hence, there is an "exchange" of tissue. • Although cellular differentiation is active in both morphallaxis and epimorphosis, in morphallaxis the majority of the regeneration comes from reorganization or exchange, while in epimorphosis the majority of the regeneration comes from cellular differentiation. HYDRA: MORPHALLACTIC REGENERATION (+ EPIMORPHOSIS) • When a Hydra is decapitated, the Wnt pathway is activated in the apical portion that will form a new head. • If the cut is made just below the hypostome, Wnt3 is up-regulated in the epithelial cells near the cut surface, which causes the remodeling of existing cells to form the head. No proliferation is seen in this case; hence, it is morphallactic regeneration (regeneration by cell transdifferentiation). • If the Hydra is cut at its midsection, however, the cells derived from the interstitial stem cell (neurons, nematocytes, secretory cells, and gametes) undergo apoptosis immediately below the cut site. Before dying, however, these cells produce a burst of Wnt3, which activates β-catenin in the interstitial cells beneath them. This β-catenin surge causes a wave of proliferation in the interstitial cells as well as remodeling in the epithelial cells. Here we have epimorphic regeneration. Canonical Wnt signaling is thus important both in normal budding and in head regeneration.
- 16. The Activation Gradients • Every portion of the hydra’s body column along the apical-basal axis is potentially able to form both a head and a foot. • The animal’s polarity, however, is coordinated by a series of morphogenetic gradients that permit the head to form only at one place and the basal disc to form only at another. • Evidence for such gradients was first obtained from grafting experiments begun by Ethel Browne in the early 1900s. When hypostome tissue from one hydra is transplanted into the middle of another hydra, the transplanted tissue forms a new apical-basal axis, with the hypostome extending outward. When a basal disc is grafted to the middle of a host hydra, a new axis also forms, but with the opposite polarity, extending a basal disc. When tissues from both ends are transplanted simultaneously into the middle of a host, either no new axis is formed or the new axis has little polarity. • These experiments have been interpreted to indicate the existence of a head activation gradient (highest at the hypostome) and a foot activation gradient (highest at the basal disc). • The higher the level of head activator in the donor tissue, the greater the percentage of implants that will induce the formation of new heads. The head activation factor is concentrated in the hypostome and decreases linearly toward the basal disc. Hypostome is the Organizer • When transplanted, (1) the hypostome can induce host tissue to form a second body axis; (2) the hypostome produces the head activation signal; (3) the hypostome is the only “self-differentiating” region of the hydra; and (4) the hypostome also produces a “head inhibition signal” that suppresses the formation of new organizing centers.
- 17. • Only the hypostome region could alter the fates of the trunk cells and cause them to become head cells. • Broun and Bode also found that the signal did not have to emanate from a permanent graft. Even transient contact with the hypostome region was sufficient to induce a new axis from a host hydra. In these cases, all the tissue of the new axis came from the host. A Gradient of Wnt3 is the Inducer • The major head inducer of the hypostome organizer is a set of Wnt proteins acting through the canonical β- catenin pathway. • These Wnt proteins are seen in the apical end of the early bud, defining the hypostome region as the bud elongates. • If the Wnt signaling inhibitor GSK3 is itself inhibited throughout the body axis, ectopic tentacles form at all levels, and each piece of the trunk has the ability to stimulate the outgrowth of new buds. • Similarly, transgenic hydra made to globally misexpress the downstream Wnt effector β-catenin form ectopic buds all along the body axis and even on top of newly formed ectopic buds. • When the hypostome is brought into contact with the trunk of an adult hydra, it induces expression of the Brachyury gene in a Wnt-dependent manner—just as vertebrate organizers do—even though hydras lack mesoderm. These results strongly indicate that Wnt proteins (in particular Wnt3) function as the head organizer during normal hydra development, but do they function similarly during regeneration?
- 19. Head Inhibition Gradients • If any region of the Hydra body column is capable of forming a head, how is head formation restricted to a specific location? • In 1926, Rand and colleagues showed that normal regeneration of the hypostome is inhibited when an intact hypostome is grafted adjacent to the amputation site. • Moreover, if a graft of sub-hypostomal tissue (where there is a relatively high concentration of head activator) is placed in the same region of a host hydra, no secondary axis forms. The host head appears to make an inhibitor that prevents the grafted tissue from forming a head and secondary axis. Supporting this hypothesis is the fact that if sub-hypostomal tissue is grafted onto a decapitated host hydra, a second axis does form. A gradient of this inhibitor appears to extend from the head down the body column and can be measured by grafting sub- hypostomal tissue into various regions along the trunks of host hydras. This tissue will not produce a head when implanted into the apical area of an intact host hydra, but it will form a head if placed lower on the host. The head inhibitor remains unknown, but it appears to be labile, with a half-life of only 2–3 hours. • It is thought that the head inhibitor and the head activator (Wnts) are both made in the hypostome, but that the head inhibition gradient falls off more rapidly than the head activator gradient. • The place where the head activator is uninhibited by the head inhibitor becomes the budding zone.
- 20. • Head formation at the base appears to be prevented by the production of another substance, a foot activator. • The inhibition gradients for the head and the foot may be important in determining where and when a bud can form. • In young adult hydras, the gradients of head and foot inhibitors appear to block bud formation. However, as the hydra grows, the sources of these labile substances grow farther apart, creating a region of tissue about two-thirds down the trunk where levels of both inhibitors are minimal. This region is where the bud forms. • Certain hydra mutants have defects in their ability to form buds, and these defects can be explained by alterations of the inhibition gradients. The L4 mutant of Hydra magnipapillata, for instance, forms buds very slowly, and does so only after reaching a size about twice as long as wild-type individuals. The amount of head inhibitor in these mutants was found to be much greater than in wild-type individuals. • Several small peptides have been found to activate foot formation, and researchers are beginning to sort out the mechanisms by which these proteins arise and function. • The specification of cells from the basal region through the body column may be mediated by a gradient of tyrosine kinase, however. The product of the shinguard gene is a tyrosine kinase that extends in a gradient from the ectoderm just above the basal disc through the lower region of the trunk. Buds appear to form where this gradient fades. The shinguard gene appears to be activated through the product of the manacle gene, a putative transcription factor that is expressed earlier in the basal disc ectoderm.
- 21. REGENERATION IN MAMMALS • Although mammals do not have the same level of regenerative abilities as the other organisms, they can regenerate certain structures. Mammals operate less on the principle of “start again from scratch” than from the premise that “if you can’t remake it, make it bigger.” • Mammals such as rodents and humans have been shown to regenerate the tips of their digits if the organism is young enough. First, a regeneration blastema composed of progenitor cells forms at the tip of the digit. As in salamander limb regeneration, re-specification does not occur. Instead, the new epidermis is derived from ectoderm-restricted progenitor cells, and the new bone comes from osteoblast progenitor cells. • HEART: It is interesting that heart tissue can also regenerate in mice, but only within the first week of neonatal life. After that, the ability is lost, presumably because the cardiomyocytes withdraw from the cell cycle. It is known, though, that cardiomyocytes in adult mammals will respond to a heart attack by re-entering the cell cycle, thus contributing to injury repair. • COMPENSATORY REGENERATION IN THE MAMMALIAN LIVER : *“Prometheus’ punishment”; *Today the standard assay for liver regeneration is a partial hepatectomy, where specific lobes of the liver are removed (after anesthesia is administered, unlike the fate of Prometheus), leaving the other hepatic lobes intact. Although the removed lobe does not grow back, the remaining lobes enlarge to compensate for the loss of the missing tissue. The amount of liver regenerated is equivalent to the amount of liver removed. • Such compensatory regeneration—the division of differentiated cells to recover the structure and function of an injured organ—is found in mammalian liver.
- 22. • The human liver regenerates by the proliferation of existing tissue. • Surprisingly, the regenerating liver cells do not fully dedifferentiate when they re-enter the cell cycle. No regeneration blastema is formed. Rather, mammalian liver regeneration appears to have two other lines of defense (for aiding regeneration), the first of which consists of normal, mature, adult hepatocytes. These mature cells, which are usually not dividing, are instructed to rejoin the cell cycle and proliferate until they have compensated for the missing part. The second line of defense is a population of hepatic progenitor cells that are normally quiescent (G0) but that are activated when the injury is severe and adult hepatocytes cannot regenerate well due to senescence, alcohol abuse, or disease. • In normal liver regeneration, the five types of liver cells—hepatocytes, duct cells, fat-storing (Ito) cells, endothelial cells, and Kupffer macrophages—all begin dividing to produce more of themselves. Each type retains its cellular identity, and the liver retains its ability to synthesize the liver-specific enzymes necessary for glucose regulation, toxin degradation, bile synthesis, albumin production, and other hepatic functions even as it regenerates itself. There are probably several redundant pathways that initiate liver cell proliferation and regeneration. • Global gene profiling indicates that the end result of these pathways is to down-regulate (but not totally suppress) the genes involved in the differentiated functions of liver cells while activating those genes committing the cell to mitosis.
- 23. • The liver stops growing when it reaches the appropriate size; the mechanism for how this is achieved is not yet known. • Some factor or factors in the blood appear to be establishing the size of the liver. • Huang and colleagues (2006) have proposed that these factors are bile acids that are secreted by the liver and positively regulate hepatocyte growth. • Partial hepatectomy stimulates the release of bile acids into the blood. These bile acids are received by the hepatocytes and activate the Fxr transcription factor, which promotes cell division. • Mice without functional Fxr protein cannot regenerate their livers. Therefore, bile acids (a relatively small percentage of the products secreted by the liver) appear to regulate the size of the liver, keeping it at a particular volume of cells. • Human livers regenerate more slowly than those of mice, but function is restored quickly. • In addition, mammalian livers possess a “second line” of regenerative ability. If the hepatocytes are unable to regenerate the liver sufficiently within a certain amount of time, the oval cells divide to form new hepatocytes. Oval cells are a small progenitor cell population that can produce hepatocytes and bile duct cells. They appear to be kept in reserve and are used only after the hepatocytes have attempted to heal the liver.