Download to read offline



![• Species temperature tolerance is related to the
prevailing climate when their order originated
– Stronger for exothermic species
• Strong phylogenetic structure in species
temperature tolerance
Findings so fair
"Bali Barat mangroves" byRonfromNieuwegein/ SouthMoretonOxfordshire, Netherlands / UK - flickr: Taman Nasional Bali Barat (West Bali National Park). Licensedunder Creative Commons Attribution-ShareAlike2.0via WikimediaCommons -
http://commons.wikimedia.org/wiki/File:Bali_Barat_mangroves.jpg#mediaviewer/File:Bali_Barat_mangroves.jpg
Christian Fischer [CC-BY-SA-3.0 (http://creativecommons.org/licenses/by-sa/3.0)], via Wikimedia CommonsBy Stef Maruch (kelp-forest.jpg) [CC-BY-SA-2.0 (http://creativecommons.org/licenses/by-sa/2.0)], via Wikimedia Commons](https://image.slidesharecdn.com/postdocbennett2016-160429074452/85/Postdoc-bennett-2016-7-320.jpg)


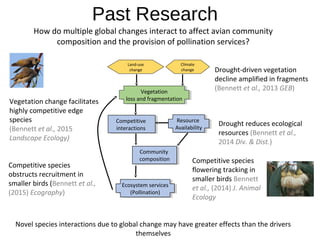
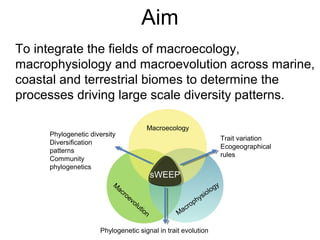
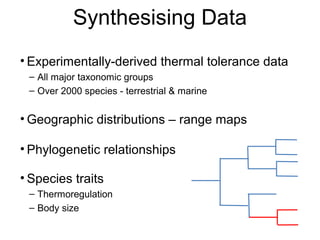
This document discusses research on how multiple global changes interact to affect avian community composition and pollination services. It summarizes past research that found vegetation decline from drought was amplified in fragmented areas, drought reduced ecological resources, and vegetation change facilitated competitive edge species. The aim of the synthesized worldwide ecology, evolution and physiology project is to integrate macroecology, macrophysiology and macroevolution across biomes to determine the processes driving large-scale diversity patterns. Some preliminary findings are that species' temperature tolerance relates to the prevailing climate when their order originated, and there is strong phylogenetic structure in temperature tolerance.



























































































![谷歌留痕技术 [ 𝙩𝙤𝙥 𝟮𝟯𝟯. 𝙘 𝙤𝙢 ]](https://cdn.slidesharecdn.com/ss_thumbnails/top233-260130174328-3833018c-thumbnail.jpg?width=640&height=640&fit=bounds)














