





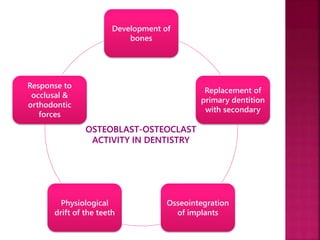


















































The document discusses the alveolar bone, which forms, supports, and protects the teeth. It has three parts: the alveolar bone proper, compact bone, and cancellous bone. Its functions include supporting teeth, attaching muscles, housing bone marrow, and storing ions. The alveolar bone is made up of water, organic components like collagen and cells, and inorganic hydroxyapatite crystals and ions. Osteoblasts form bone by synthesizing an uncalcified bone matrix and depositing hydroxyapatite crystals, while osteoclasts resorb bone by secreting acids and enzymes to dissolve the mineral and collagen phases.




































































































