Download to read offline






















![Intermediate Dominan & Muatan Mutasi
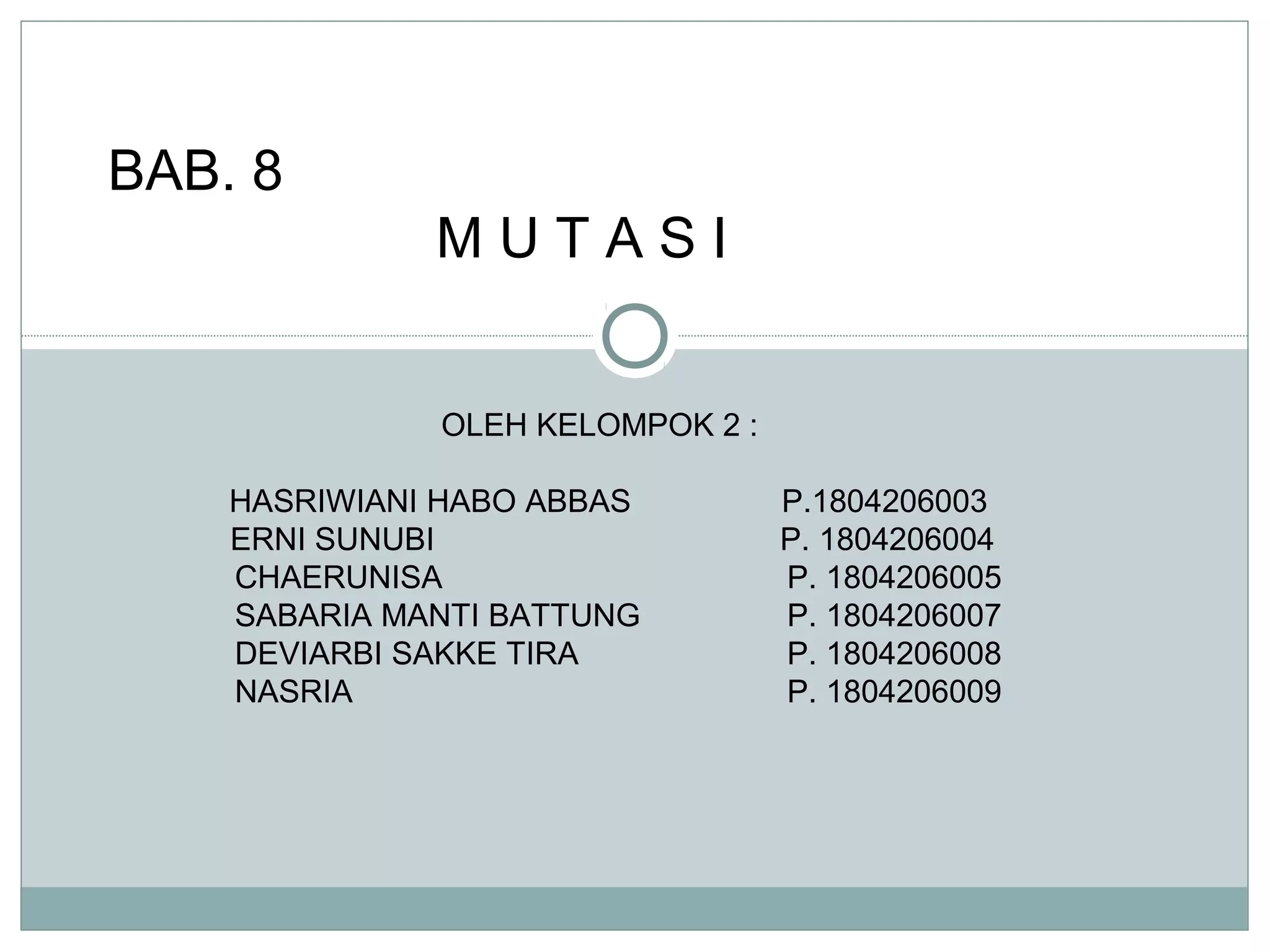
U/ memeriksa pengaruh dari level dominan, mk
diasumsikan total perubahan dlm frekuensi alel
adlh :
∆q = ∆qmu + ∆qx
Dgn level intermediate dominan dmn kondisi
(kebugaran/kesehatan) relatif dari heterozygot
adlh 1-hs,shg persamaan ini mjd :
∆q = up − spq[ h − ( 2h − 1) q ]](https://image.slidesharecdn.com/mutasi-140213215944-phpapp02/85/mutasi-sma-24-320.jpg)
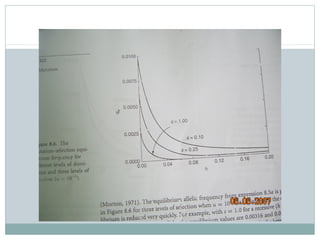
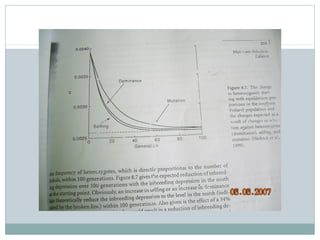





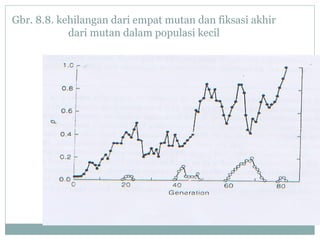


















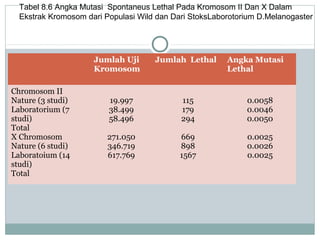





Dokumen ini membahas proses mutasi dalam populasi genetik dan pentingnya sebagai sumber variasi genetik. Proses ini dapat disebabkan oleh berbagai faktor, seperti kesalahan replikasi DNA dan pengaruh mutagen, serta memiliki peran dalam evolusi. Selain itu, analisis terhadap mutasi forward dan backward serta pengaruh seleksi terhadap frekuensi alel juga dijelaskan, menunjukkan interaksi kompleks antara mutasi dan evolusi genetik.










































































![Modul Ajar Kurikulum Berbasis Cinta (KBC) SKI Kelas 11 [modulguruku.com]](https://cdn.slidesharecdn.com/ss_thumbnails/modulajarkurikulumberbasiscintakbcskikelas11mamodulguruku-260211160930-802ab0b6-thumbnail.jpg?width=640&height=640&fit=bounds)