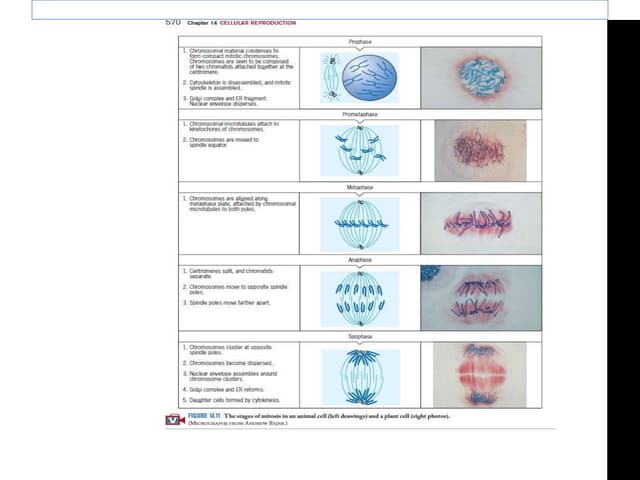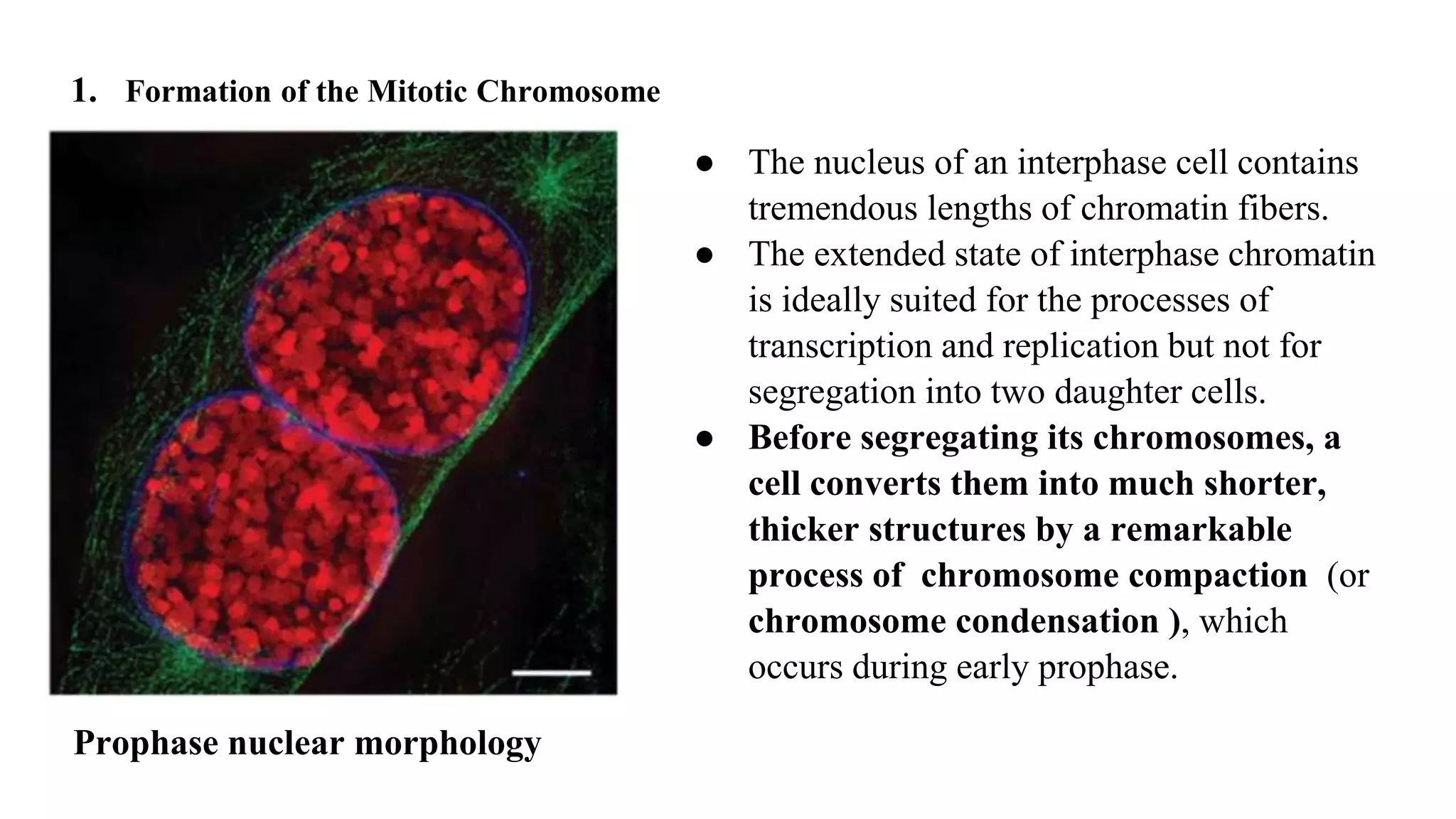


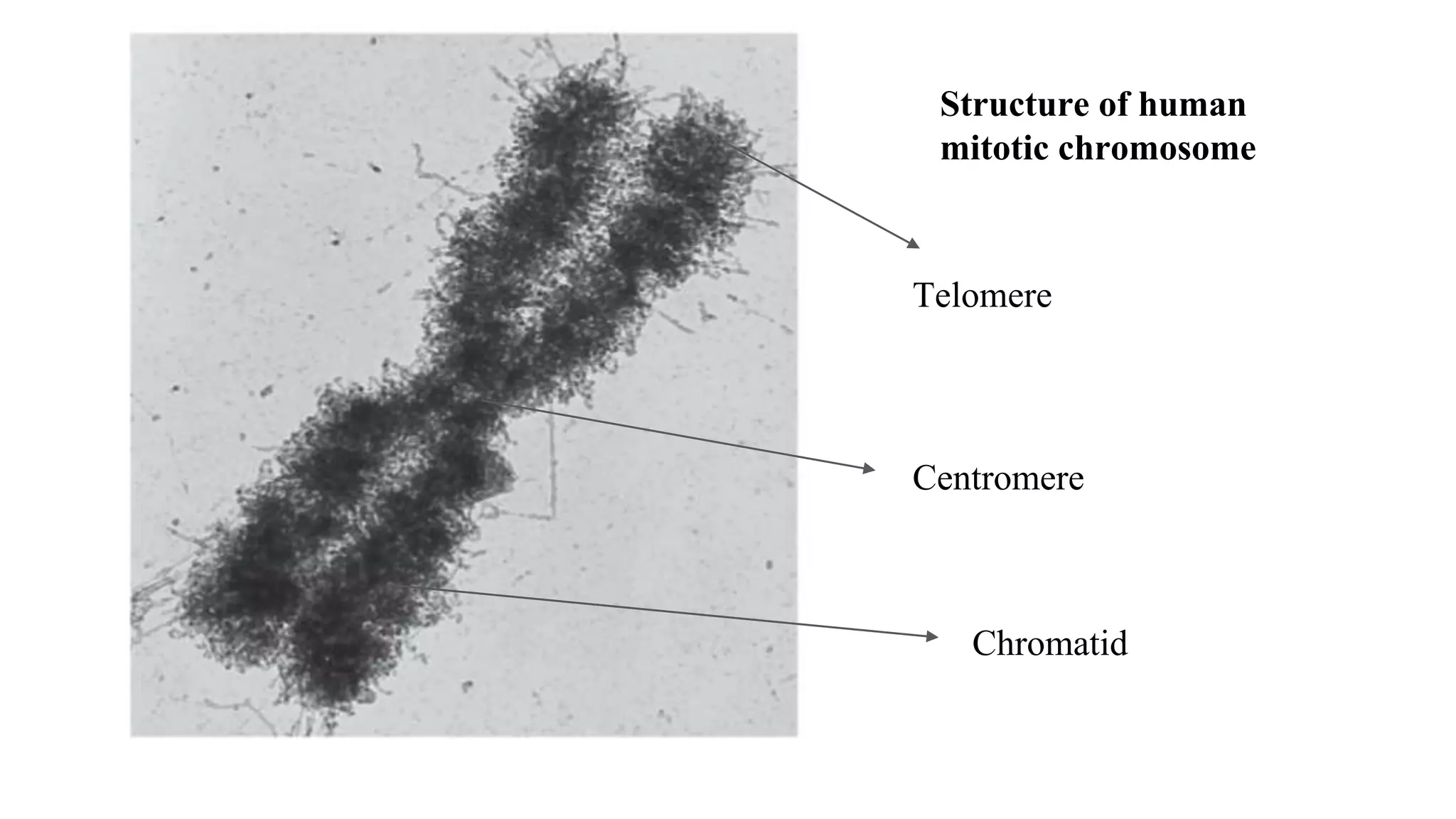





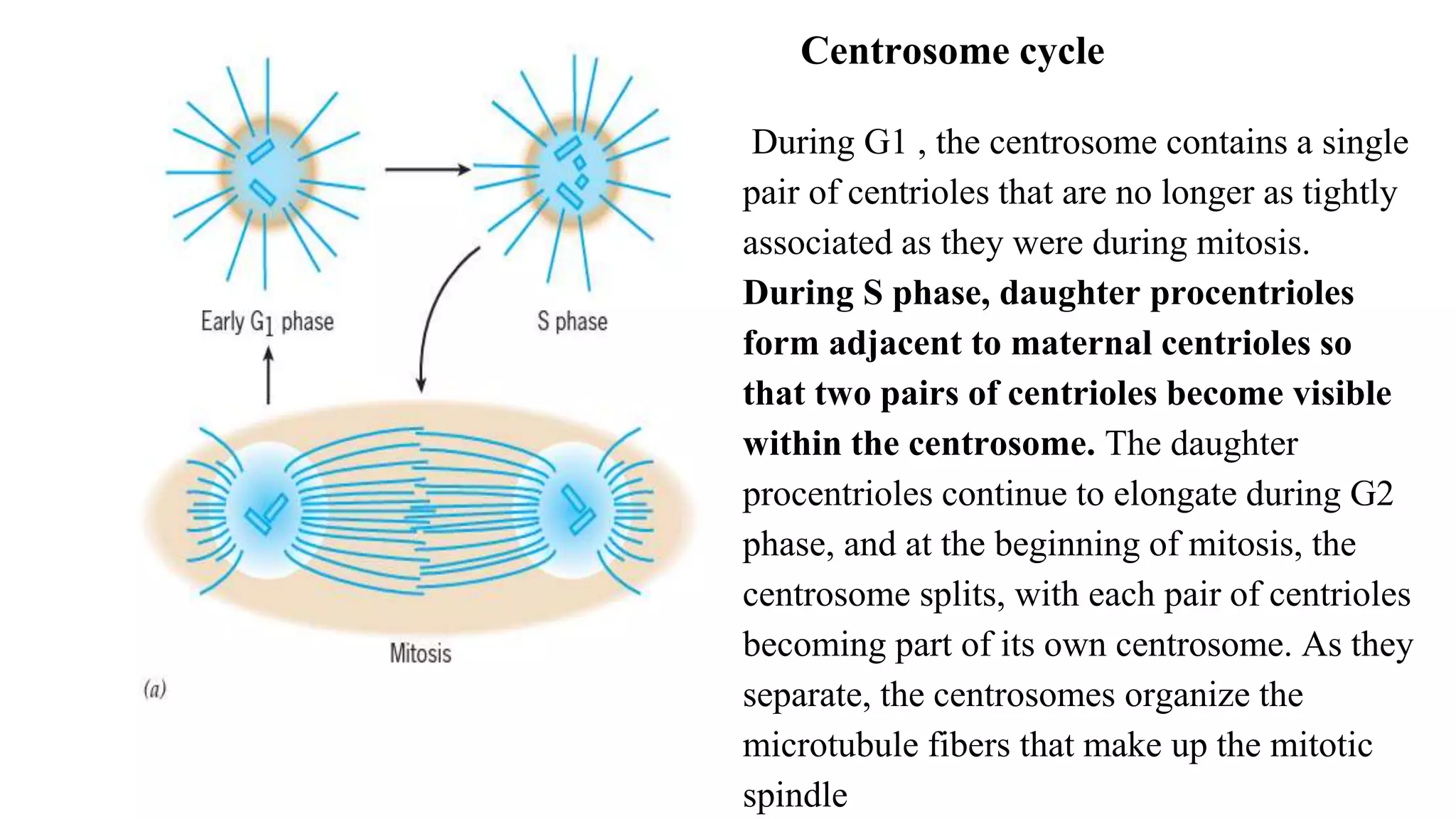





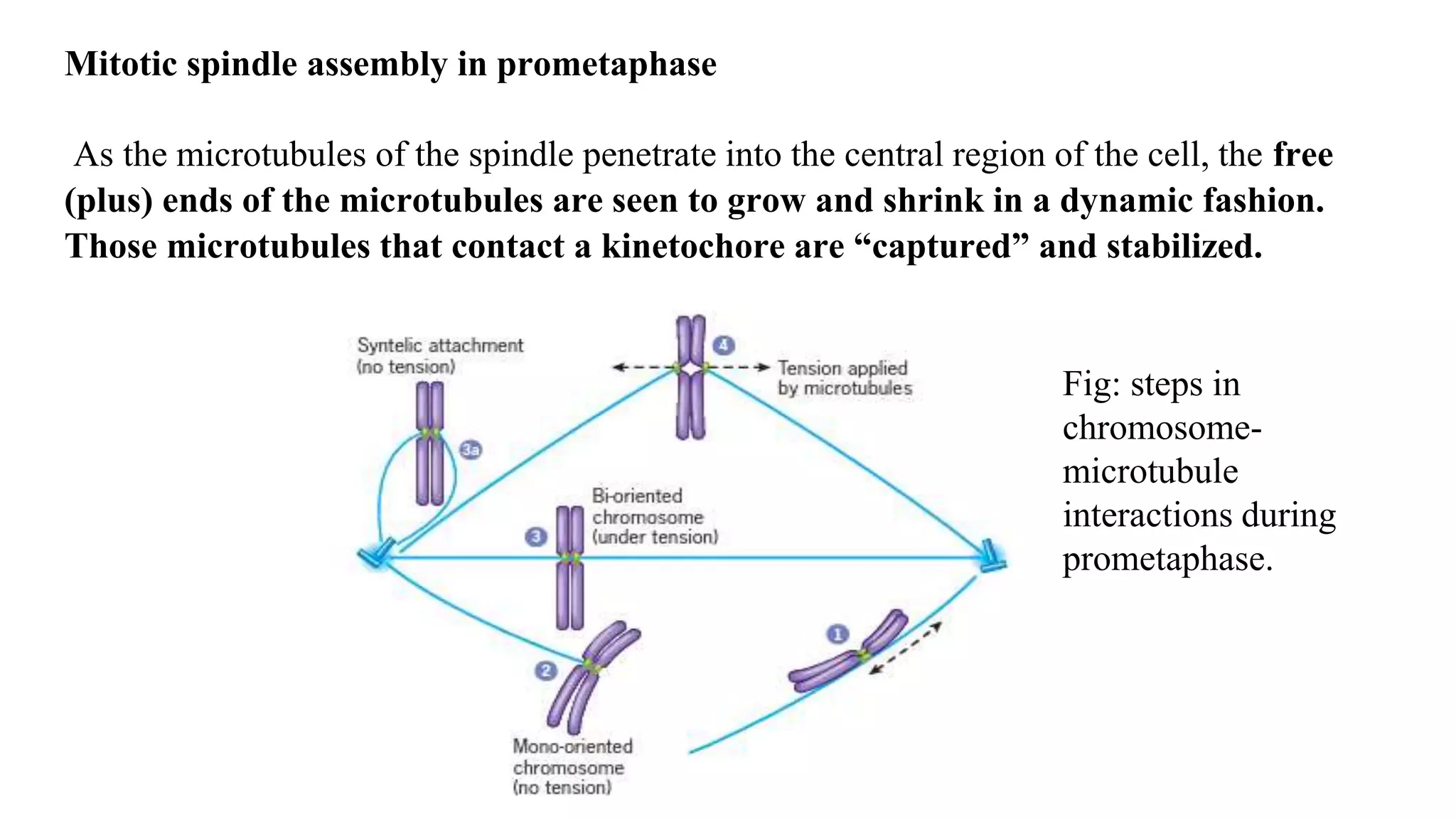




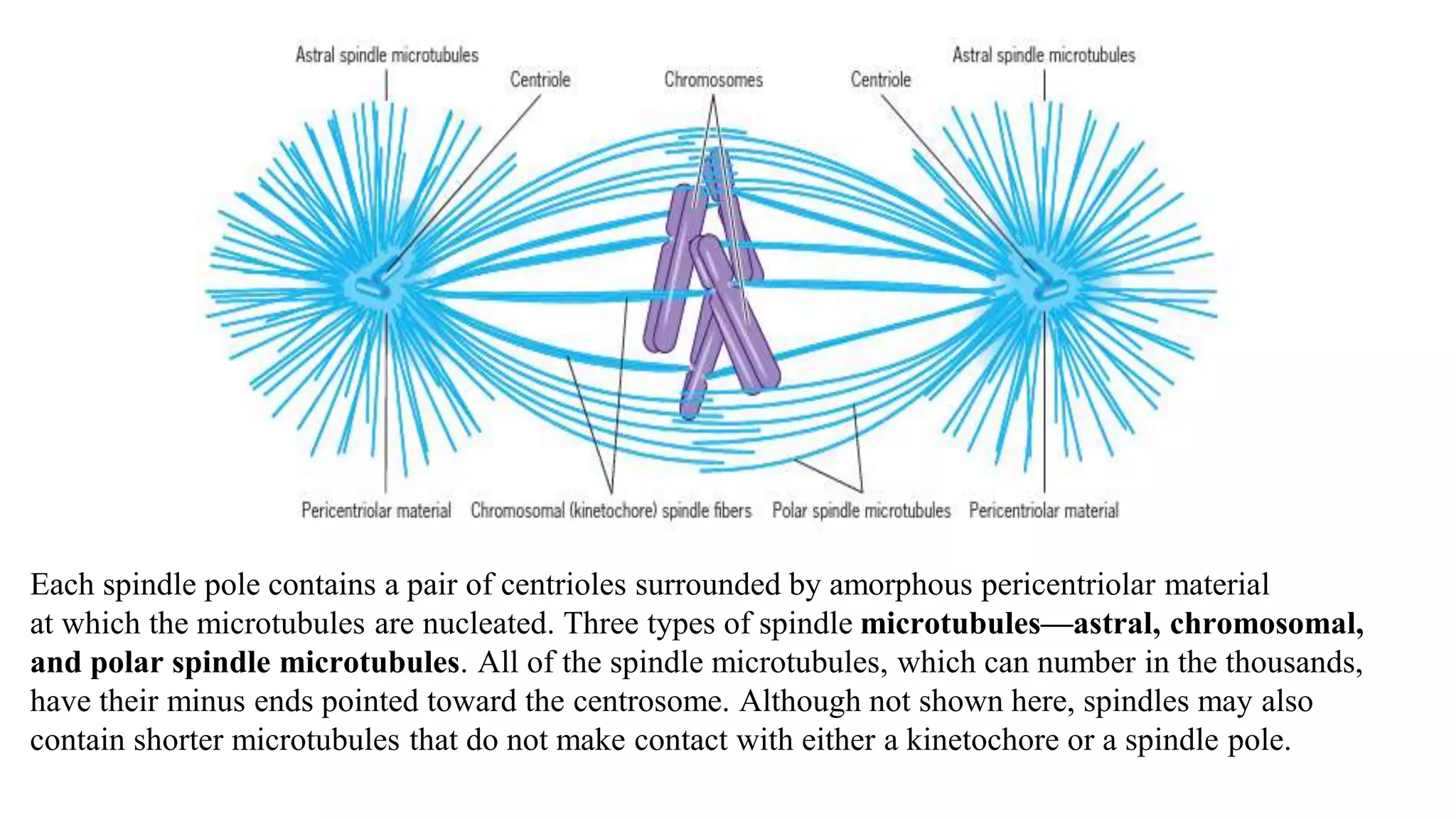

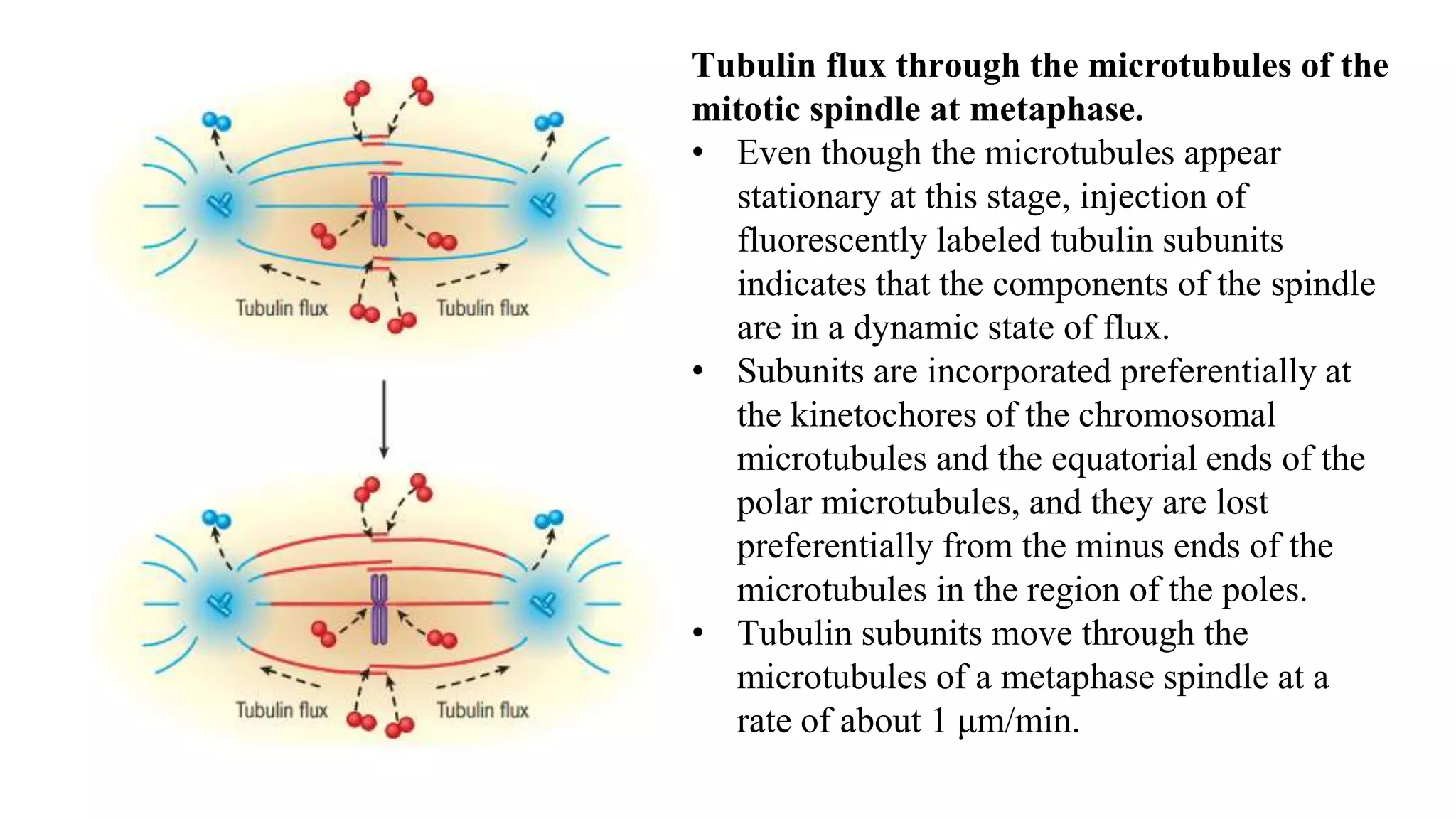



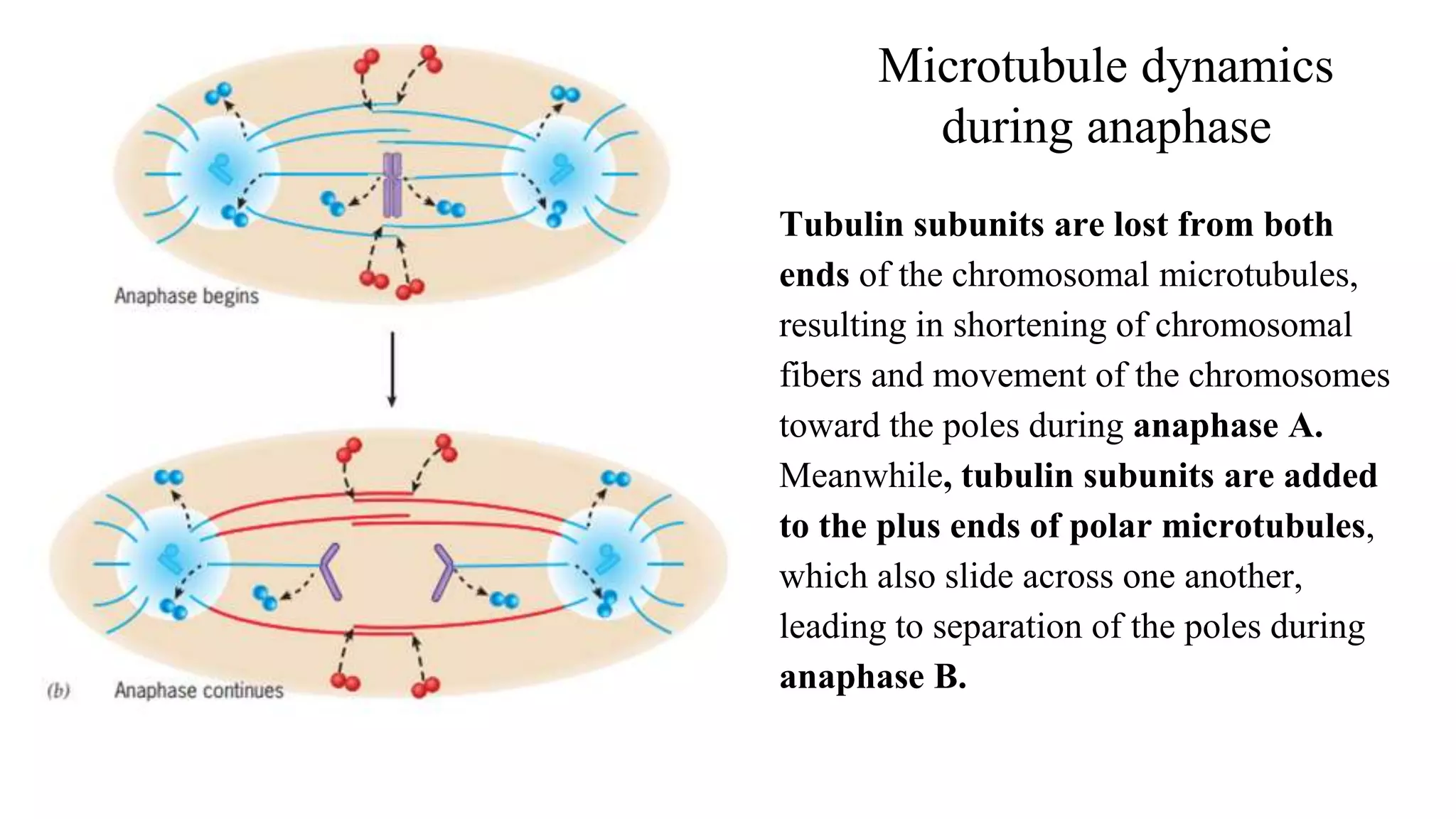








The document outlines the process of cell division, specifically mitosis, detailing its phases, the roles of various structures such as kinetochores and microtubules, and the genetic outcomes for daughter cells. It explains the intricate steps during prophase, prometaphase, metaphase, and anaphase, including chromosome compaction, nuclear envelope dissolution, spindle formation, and chromatid separation. Mitosis is essential for growth and maintenance in organisms, yielding two genetically identical daughter cells from a single parent cell.