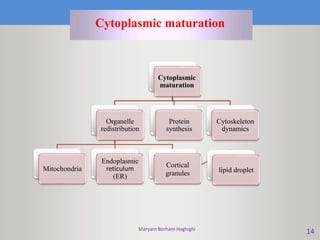
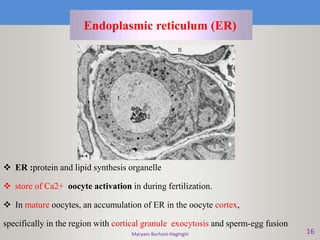
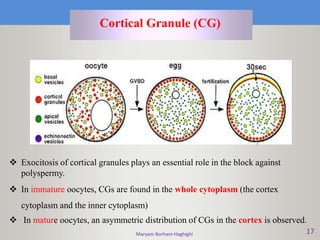
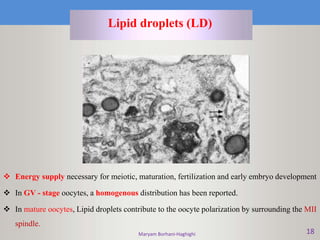
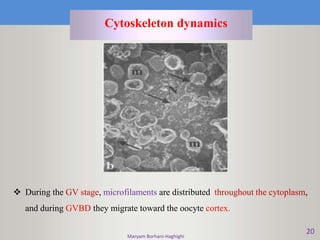
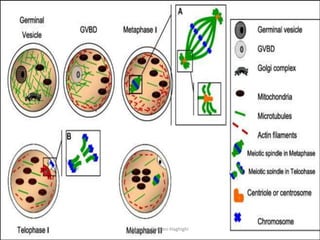
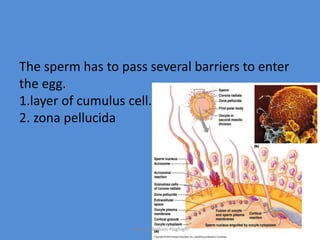
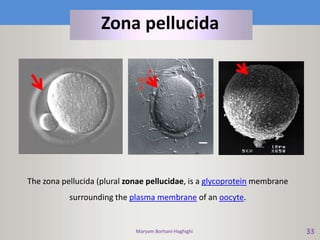
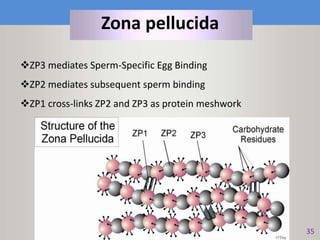
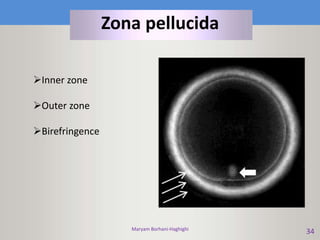
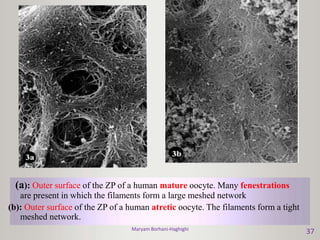
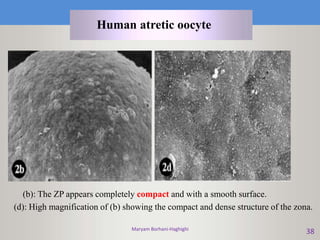
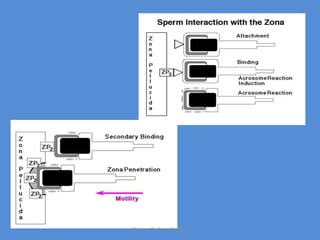
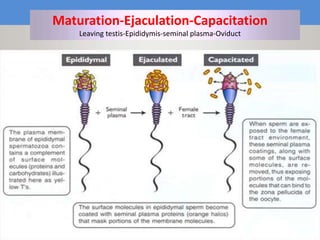
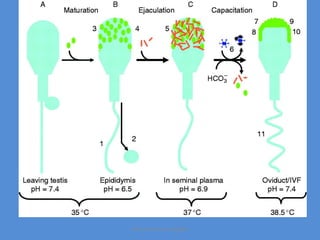
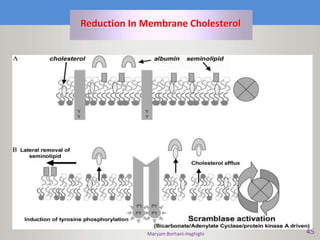
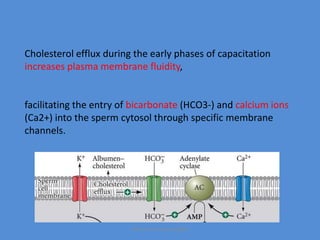
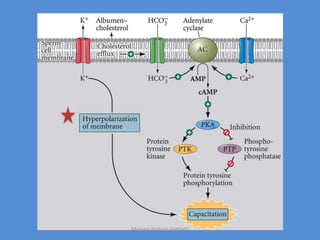
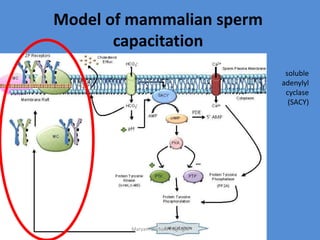
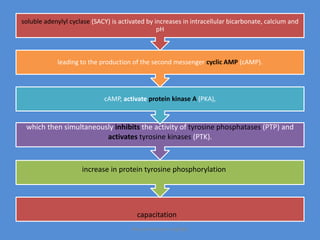
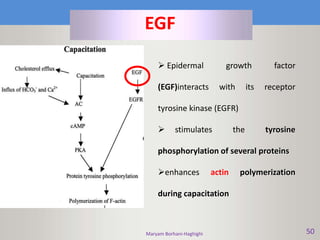
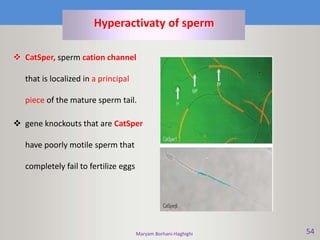
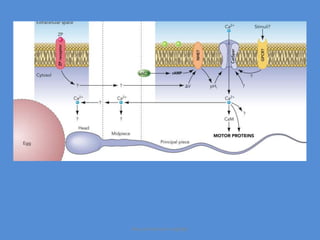
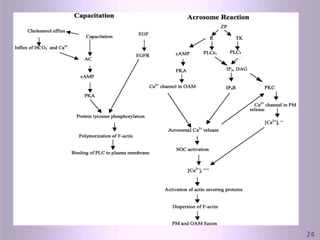
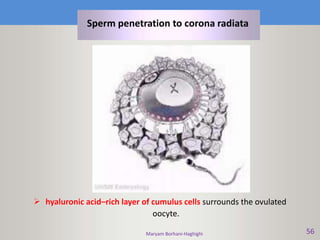
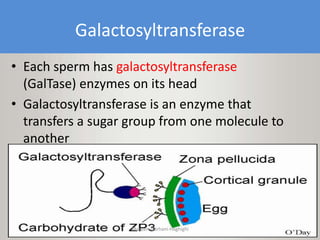
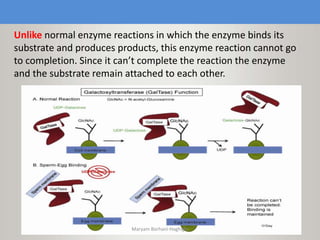
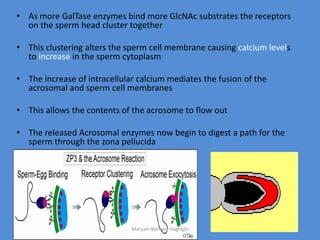
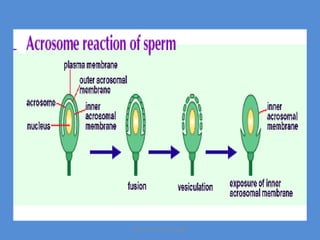
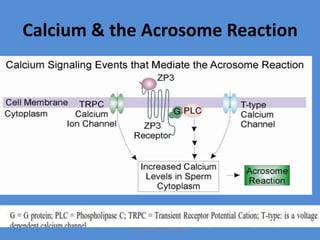
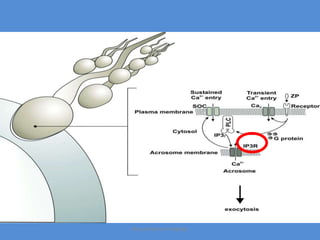
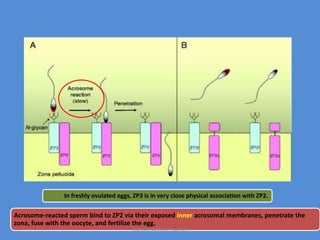
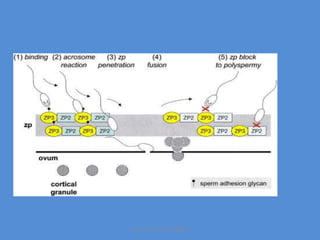
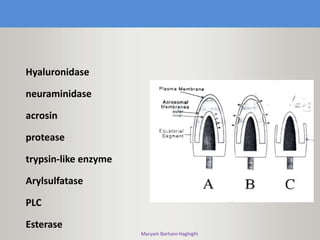
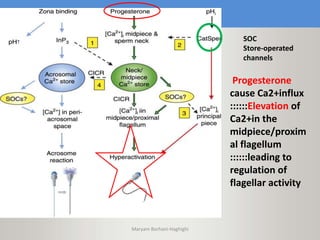
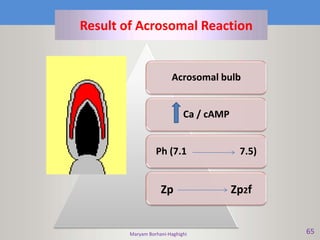
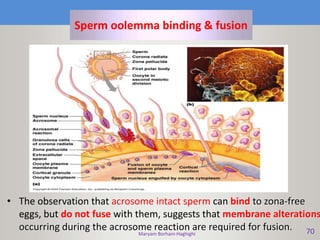
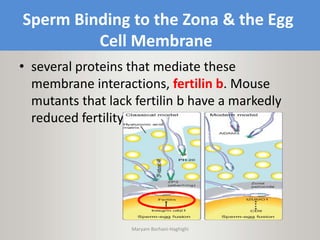
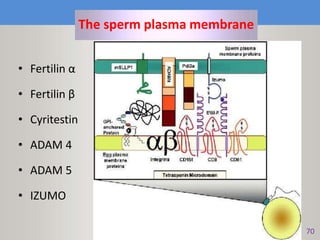
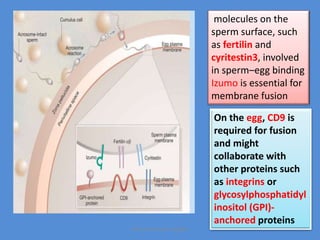
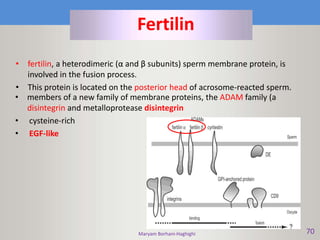
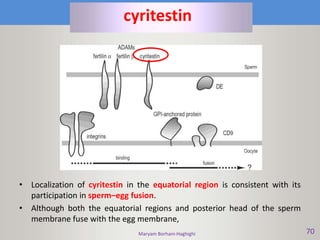
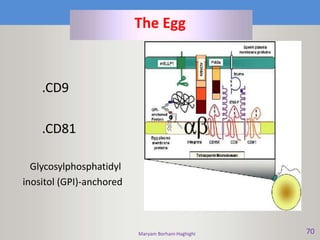
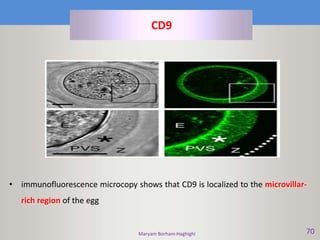
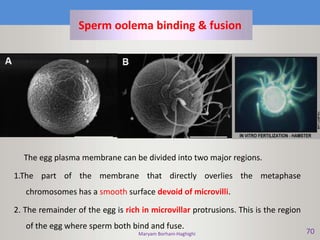
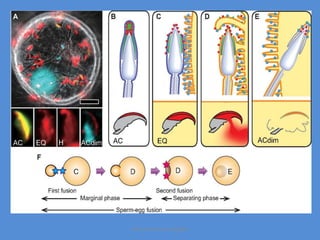
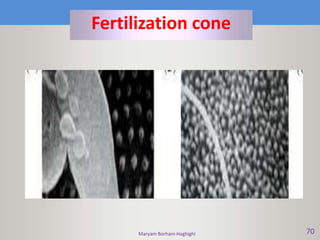
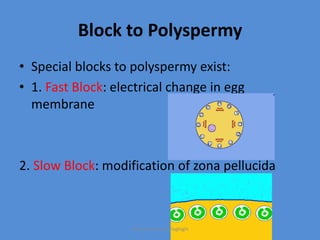
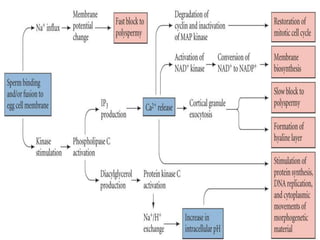
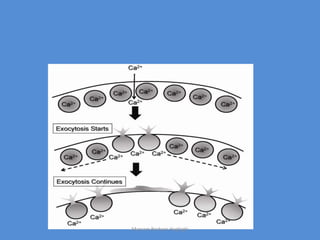
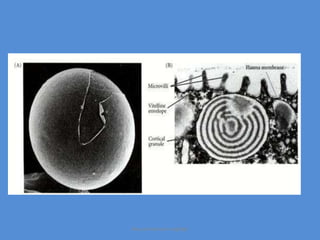
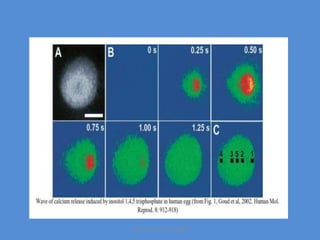
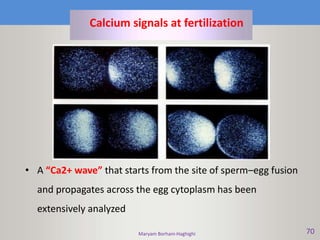
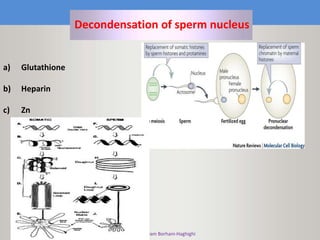
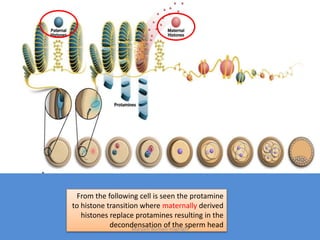
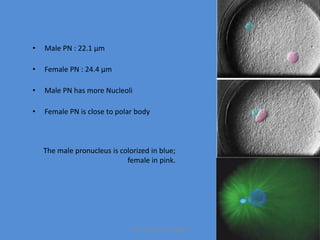
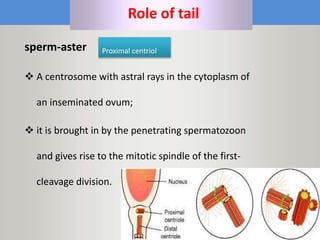
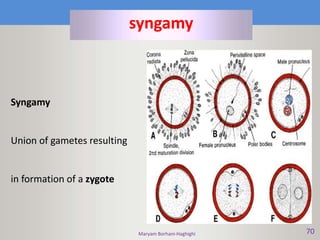
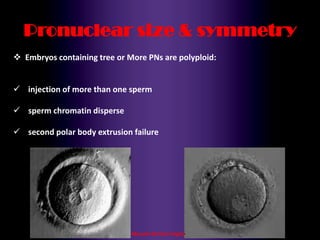
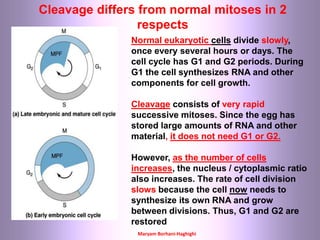
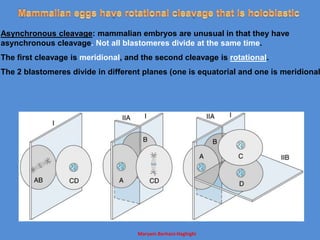
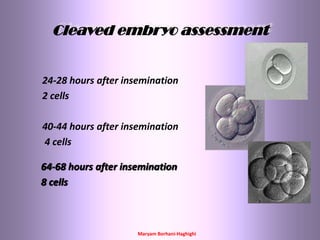
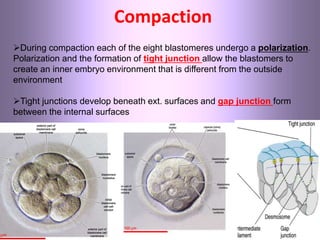
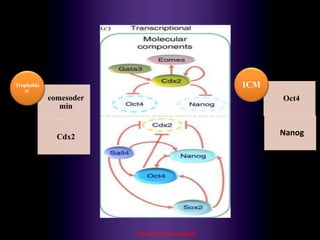
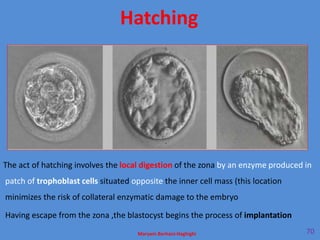
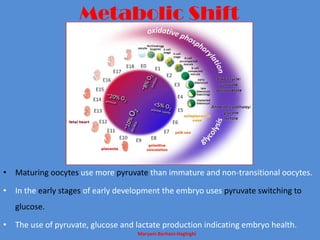
This document discusses oocyte maturation and the processes involved in fertilization. It begins with an overview of nuclear and cytoplasmic maturation during oocyte development. It then describes the structures and organelles involved in cytoplasmic maturation, including redistribution of mitochondria, endoplasmic reticulum, cortical granules, and lipid droplets. The document next explains the capacitation of sperm in the female reproductive tract. It discusses the molecular changes involved in capacitation and the associated increases in motility. The document concludes by outlining the steps of fertilization, including sperm penetration of the zona pellucida and associated acrosome reaction, binding and fusion of sperm with the oocyte membrane.





































































































![PERI-PROSTHETIC FRACTURE NAIL-PLATE CONSTRUCT [NPC].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/drarunkumardrmohamedashrafperiprostheticfrasturenail-plateconstructnpc-260209164459-7e9d15a1-thumbnail.jpg?width=640&height=640&fit=bounds)


















