Downloaded 27 times







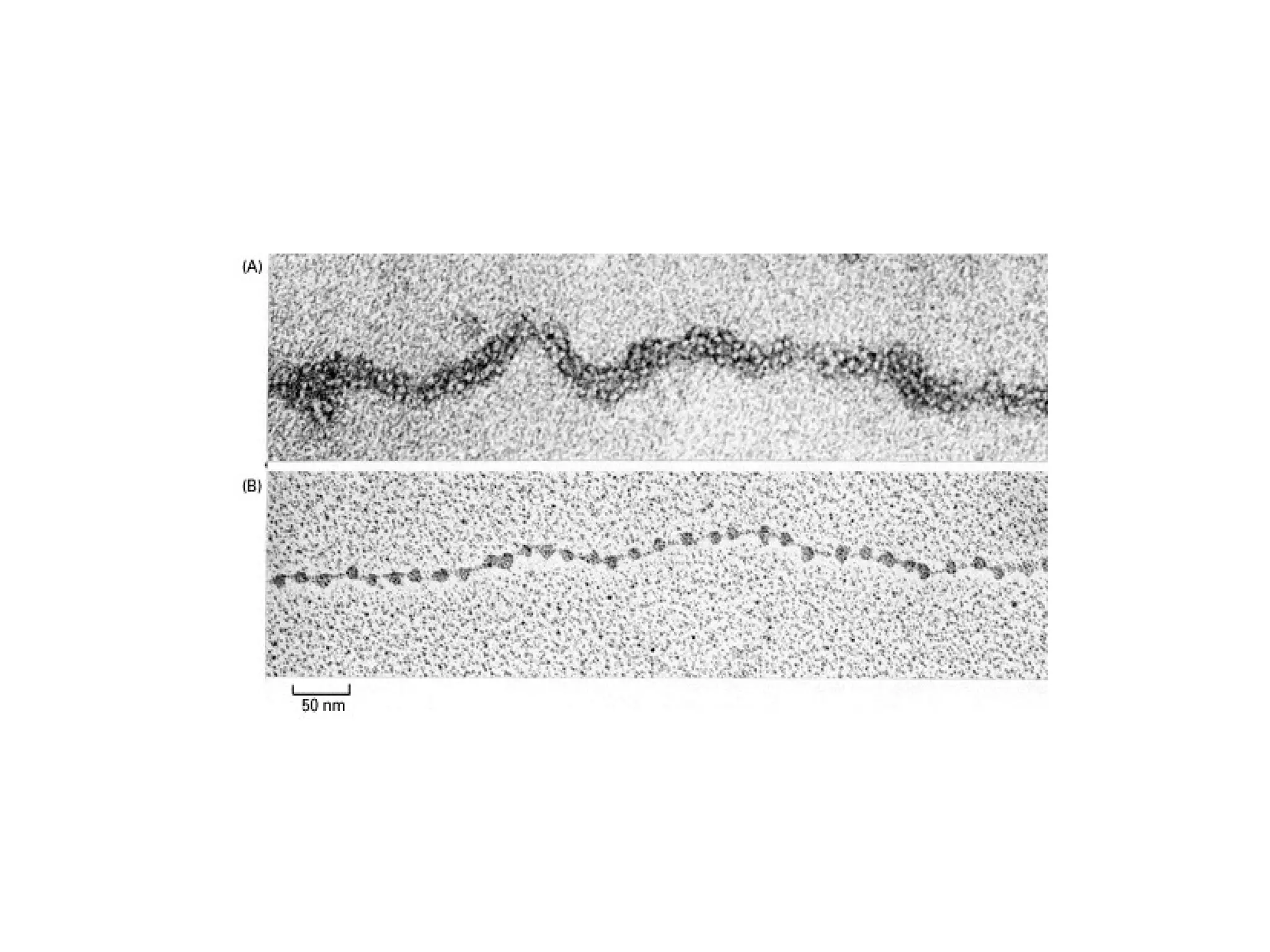












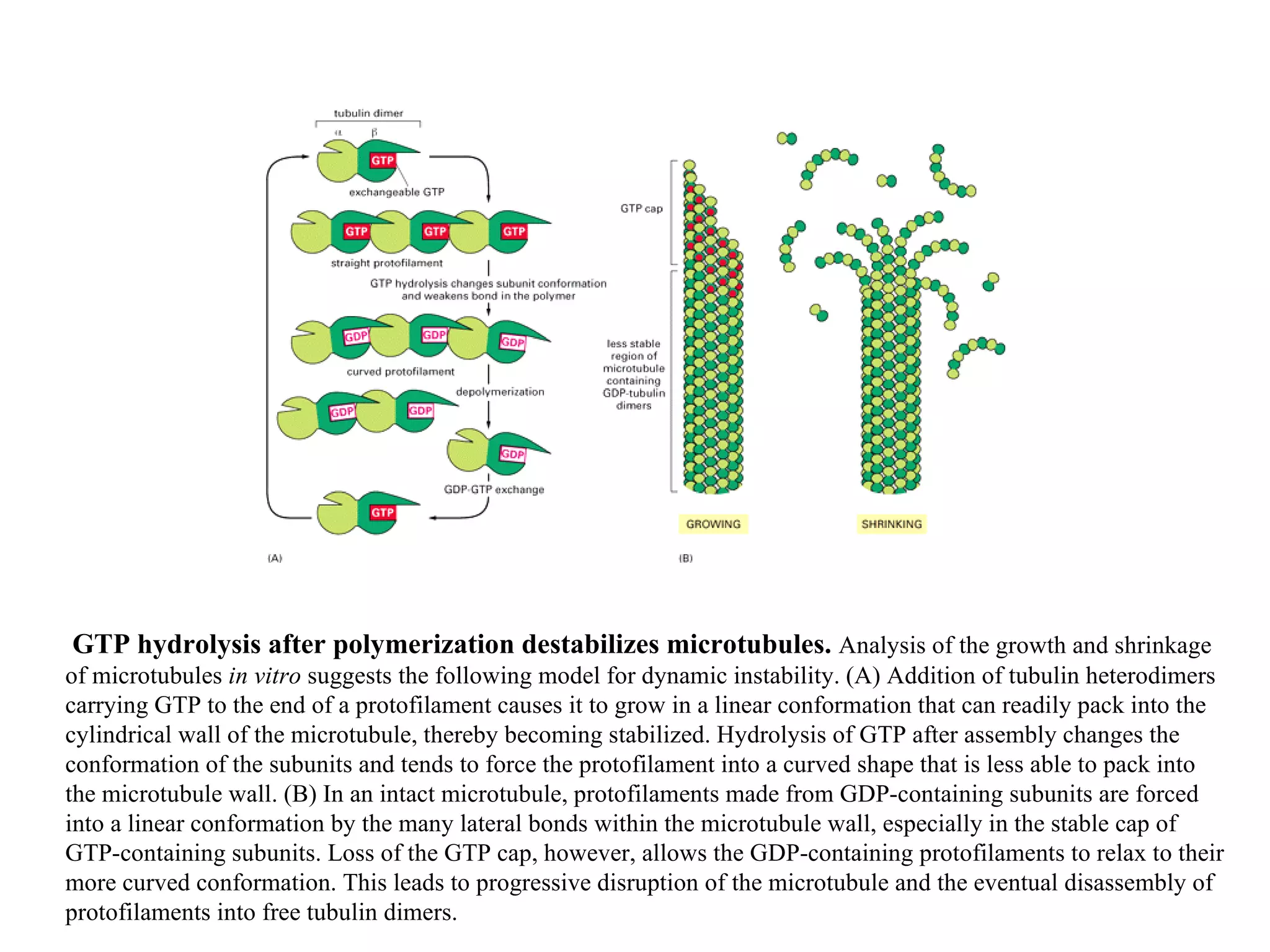
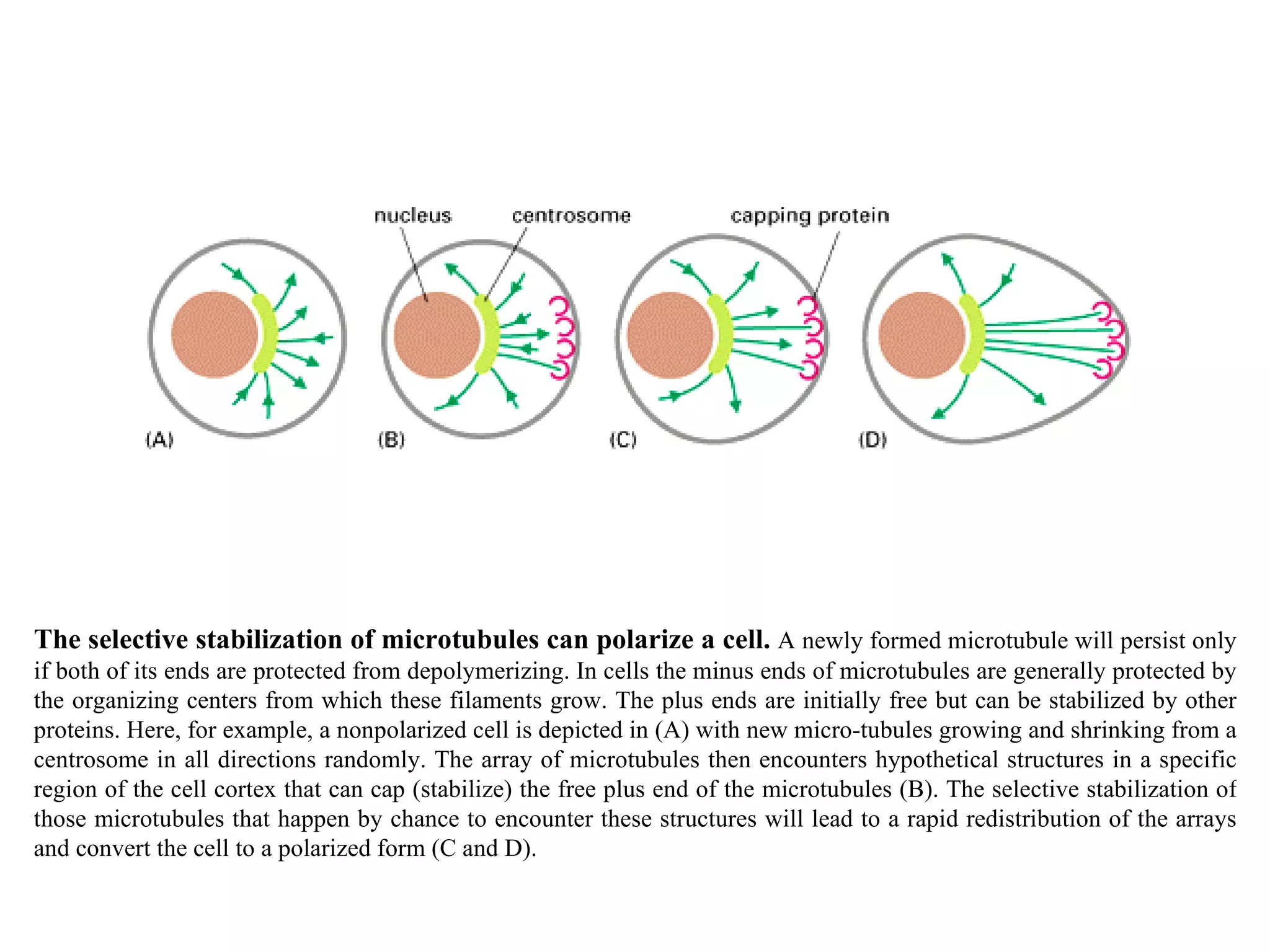

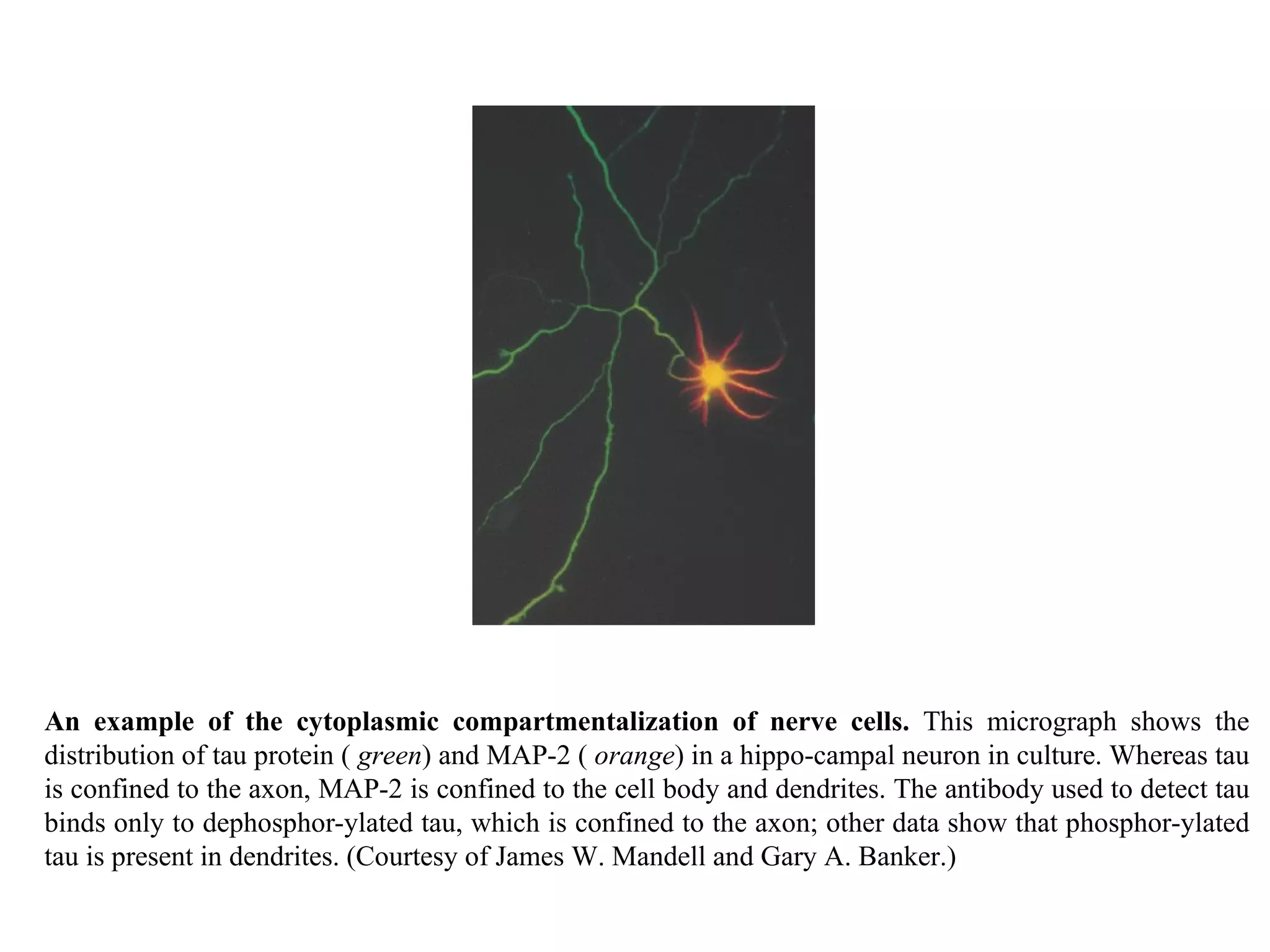
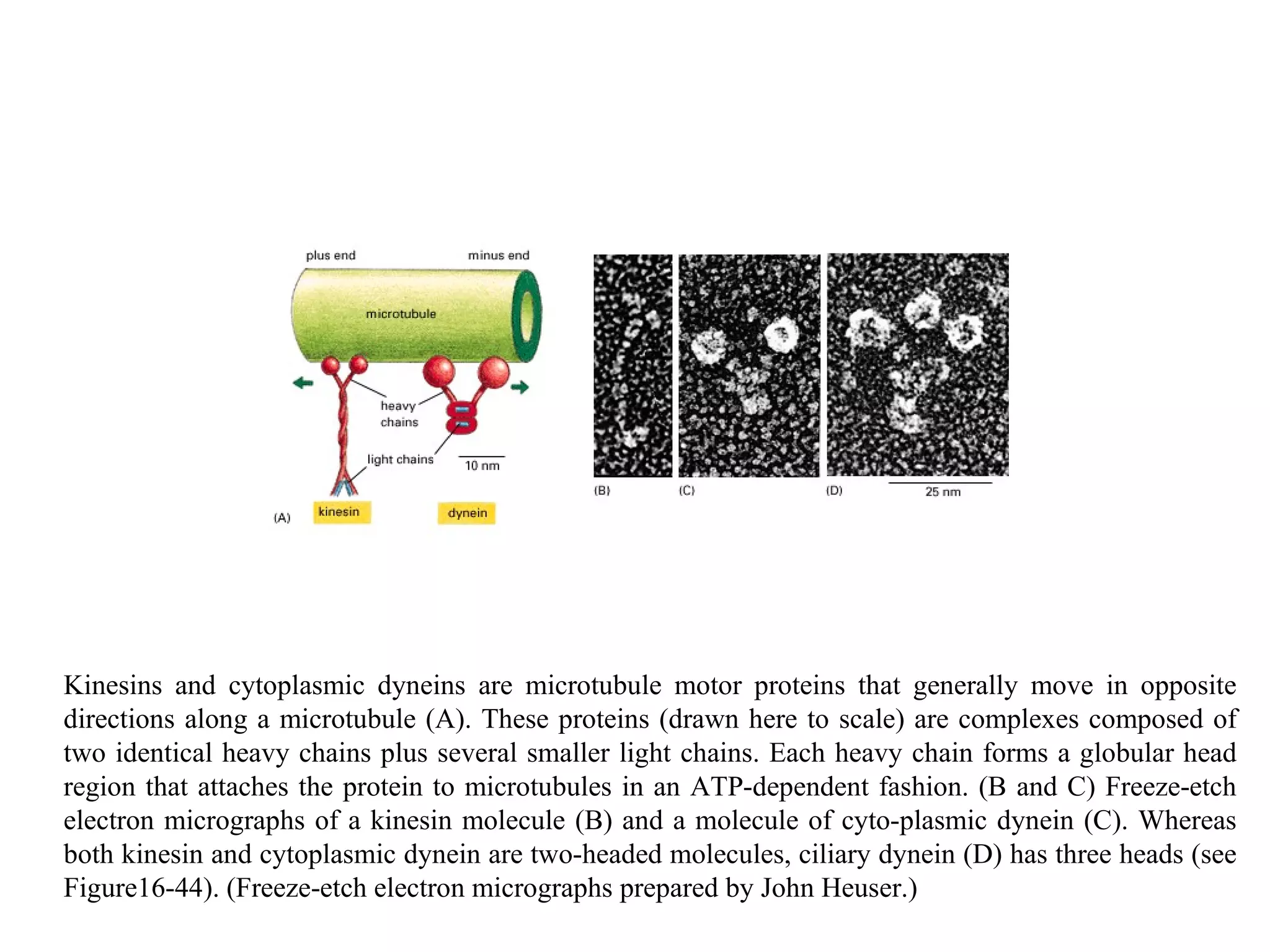



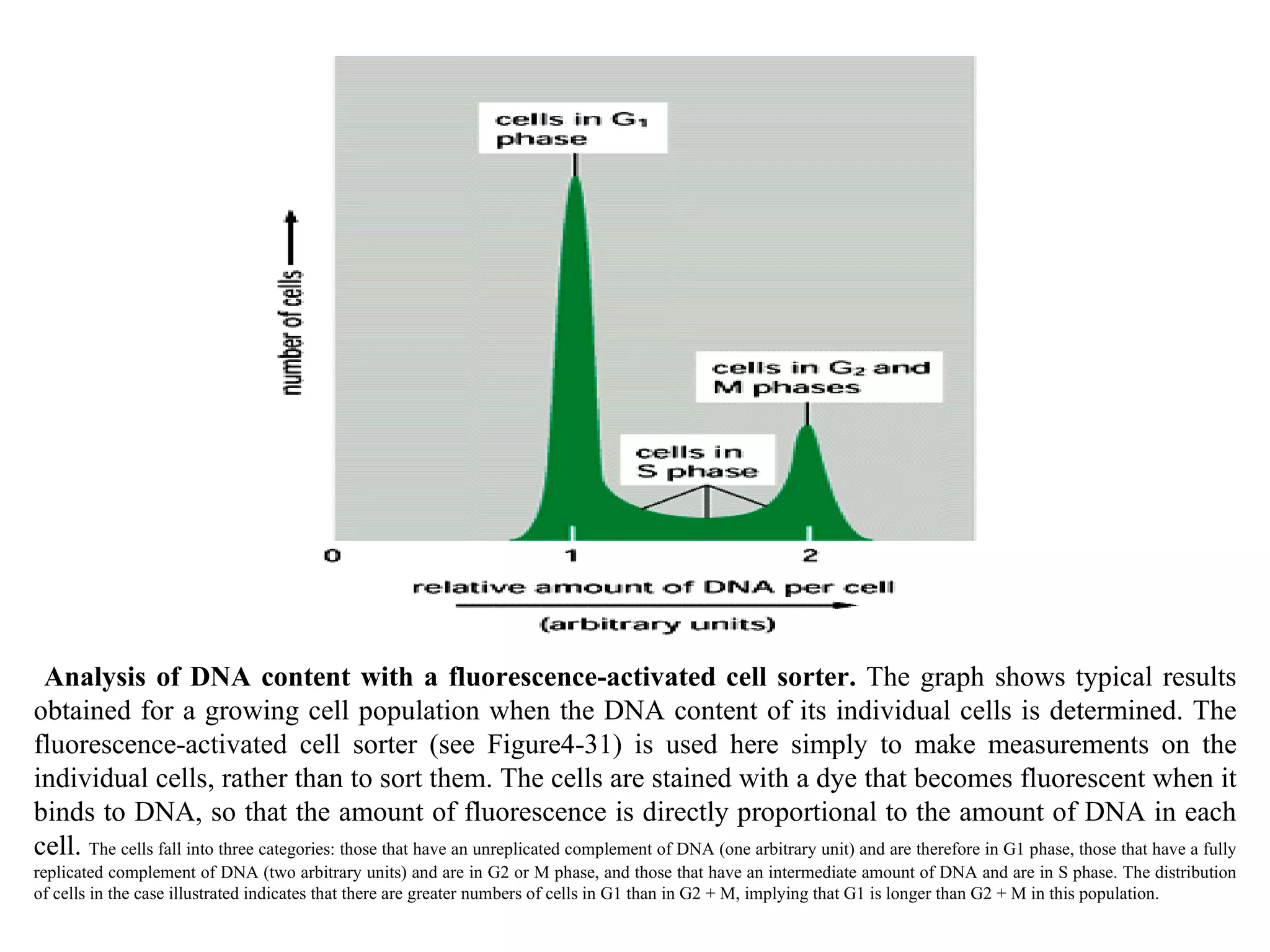
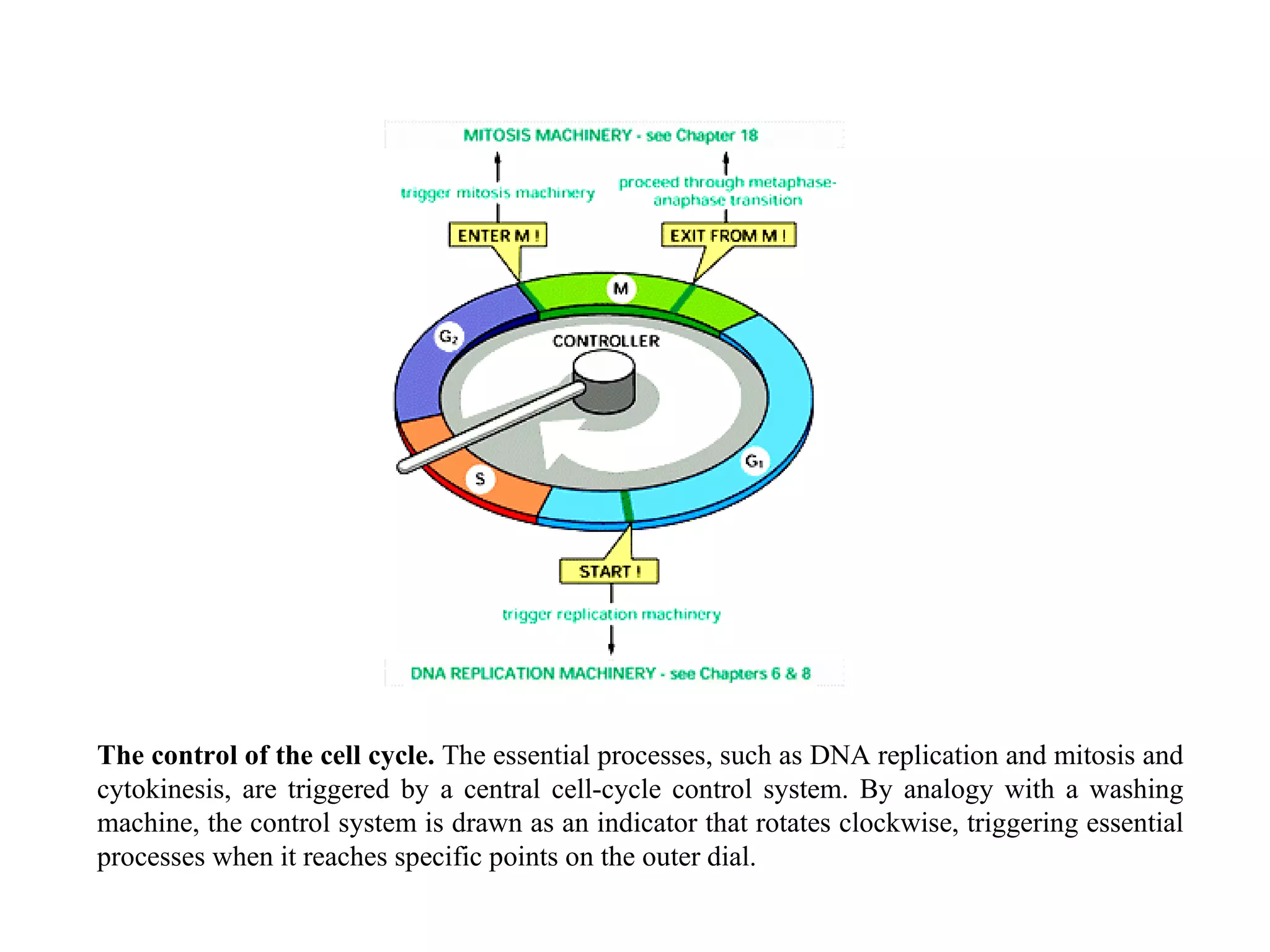
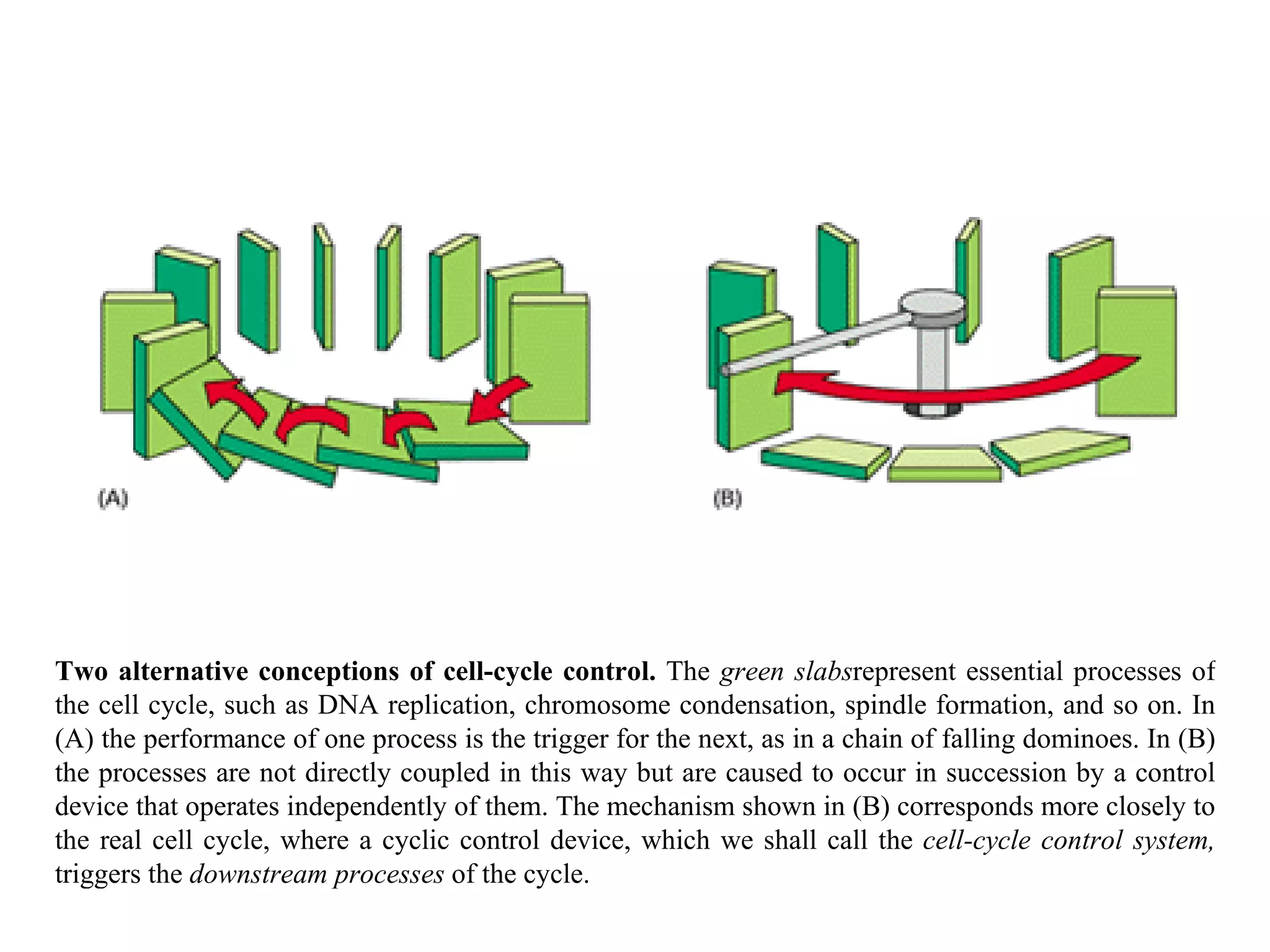
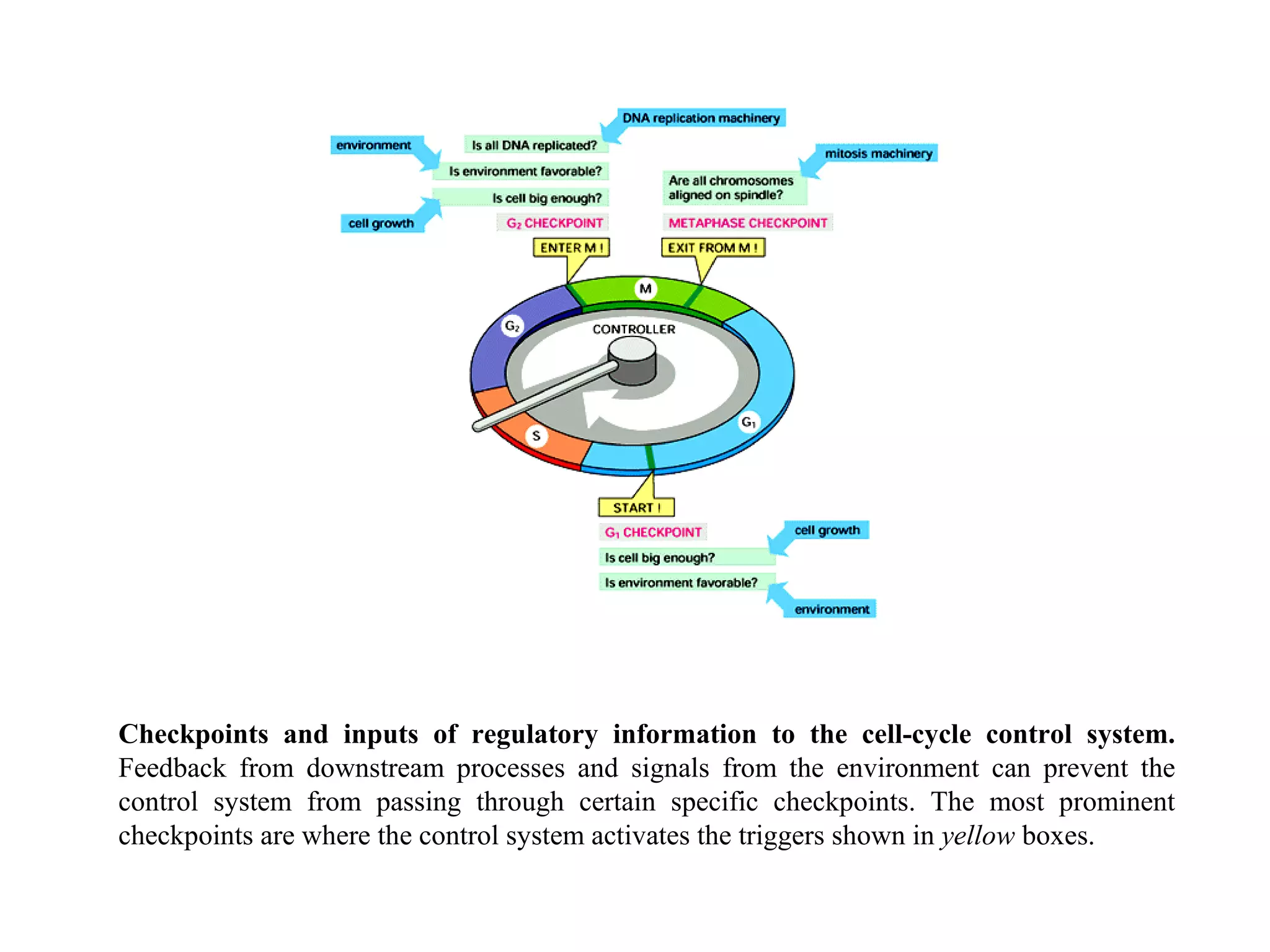
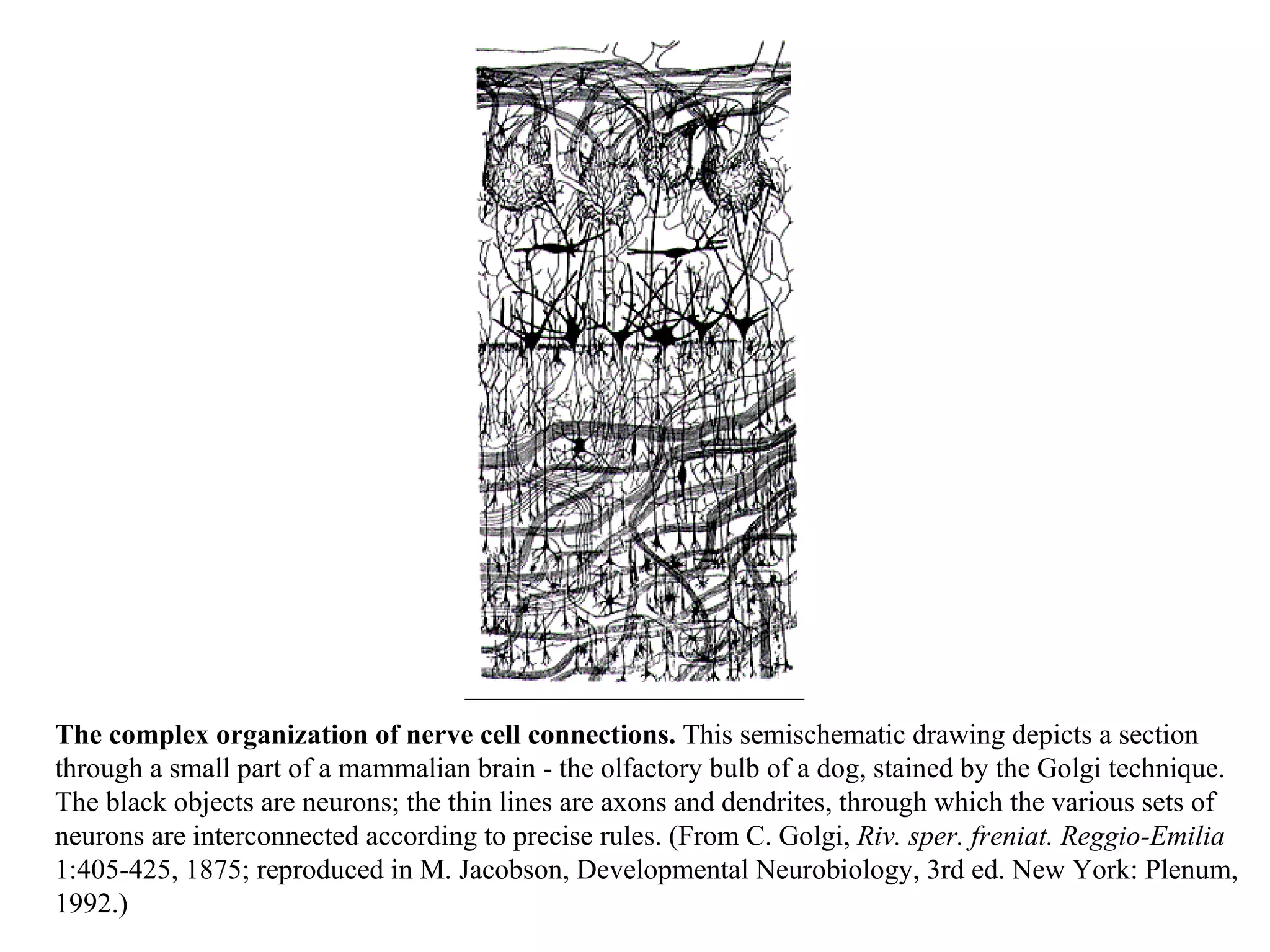
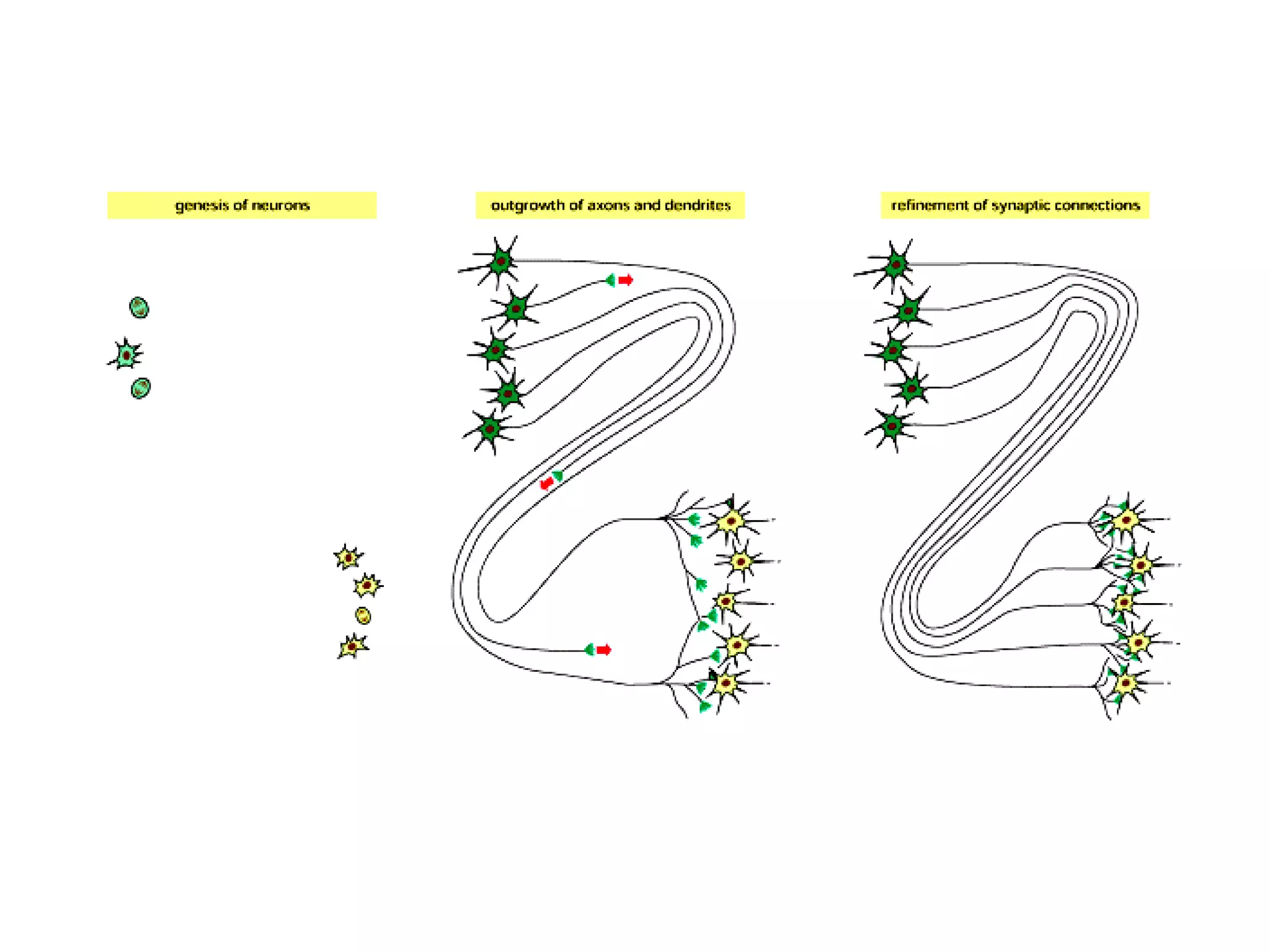
![A typical neuron of a vertebrate. The arrows indicate the direction in which signals are conveyed. The neuron shown is from the retina of a monkey. The longest and largest neurons in a human extend for about 1 million µm and have an axon diameter of 15 µm. (Drawing of neuron from B.B. Boycott in Essays on the Nervous System [R. Bellairs and E.G. Gray, eds.]. Oxford, UK: Clarendon Press, 1974.)](https://image.slidesharecdn.com/sprintfinal04-b-091203115615-phpapp02/75/Sprint-Final-09-B-94-2048.jpg)

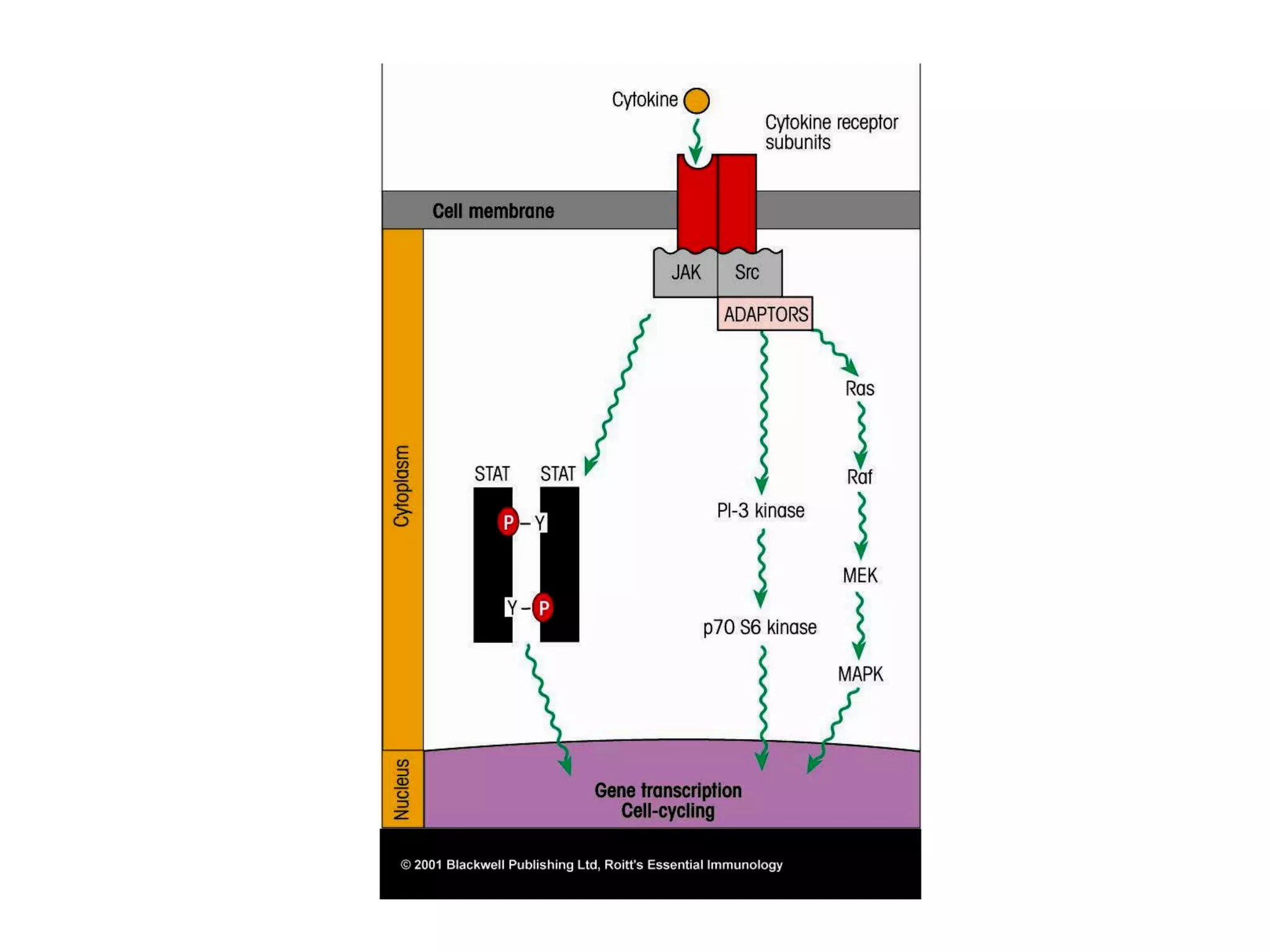
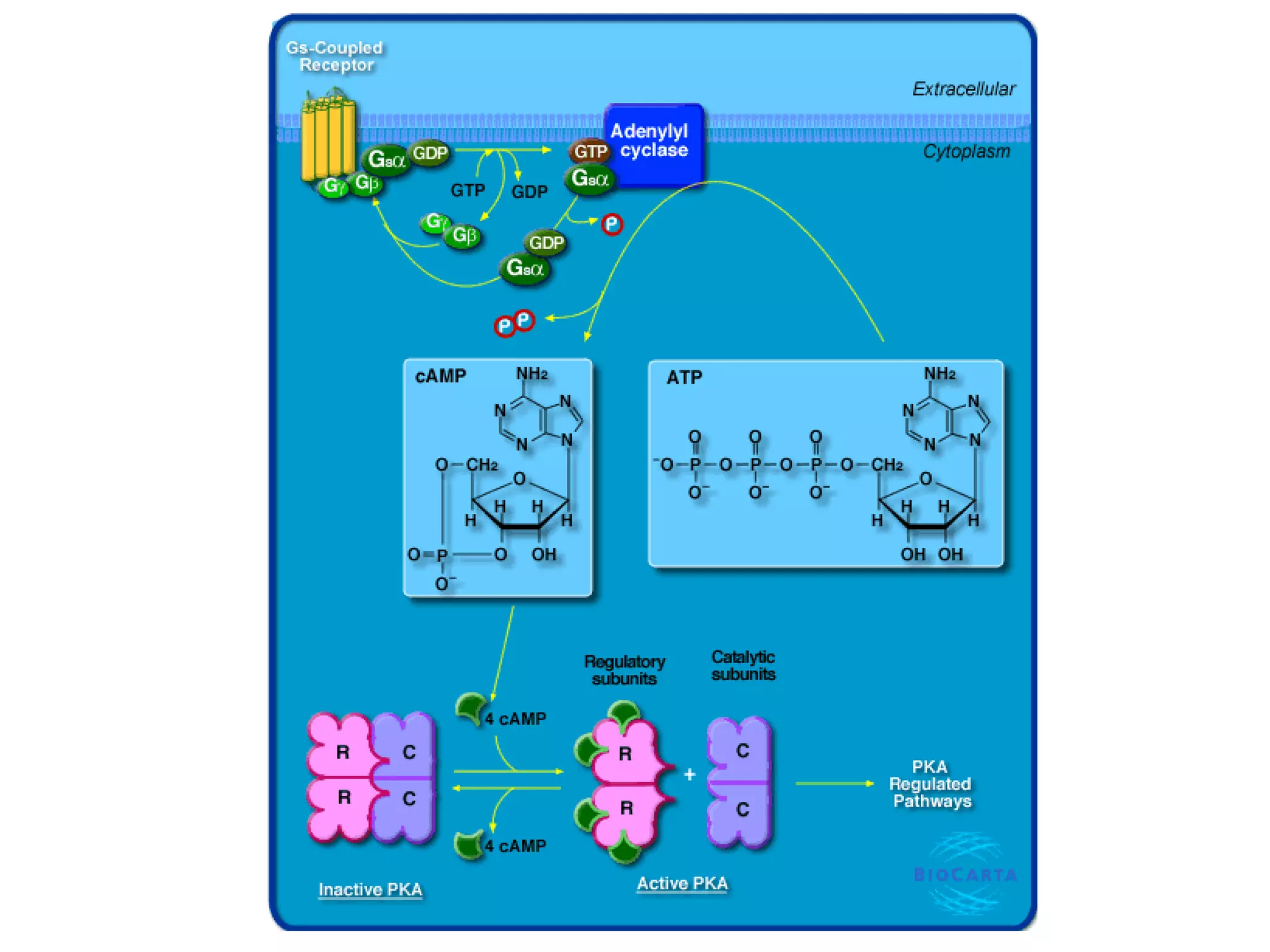

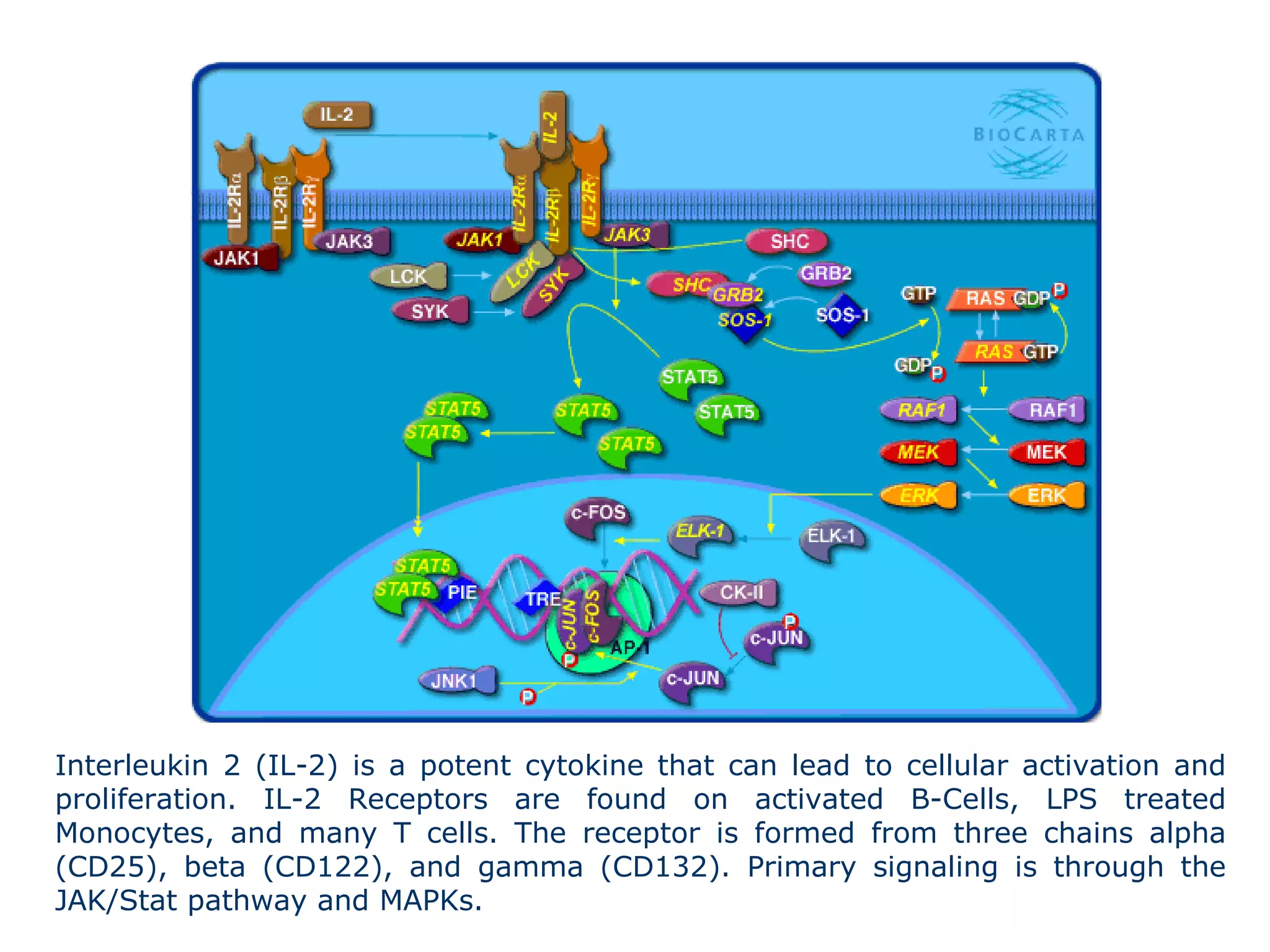
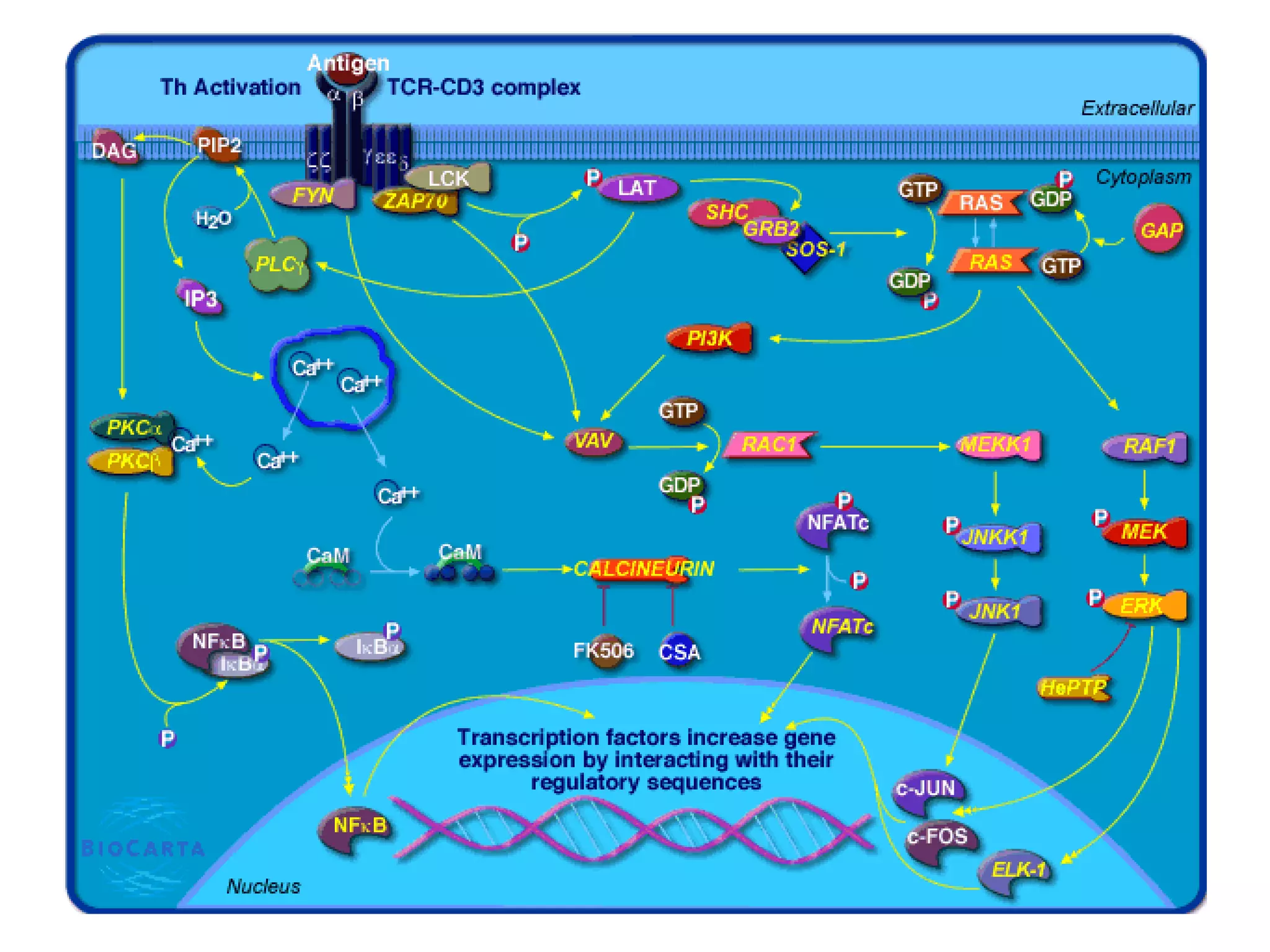


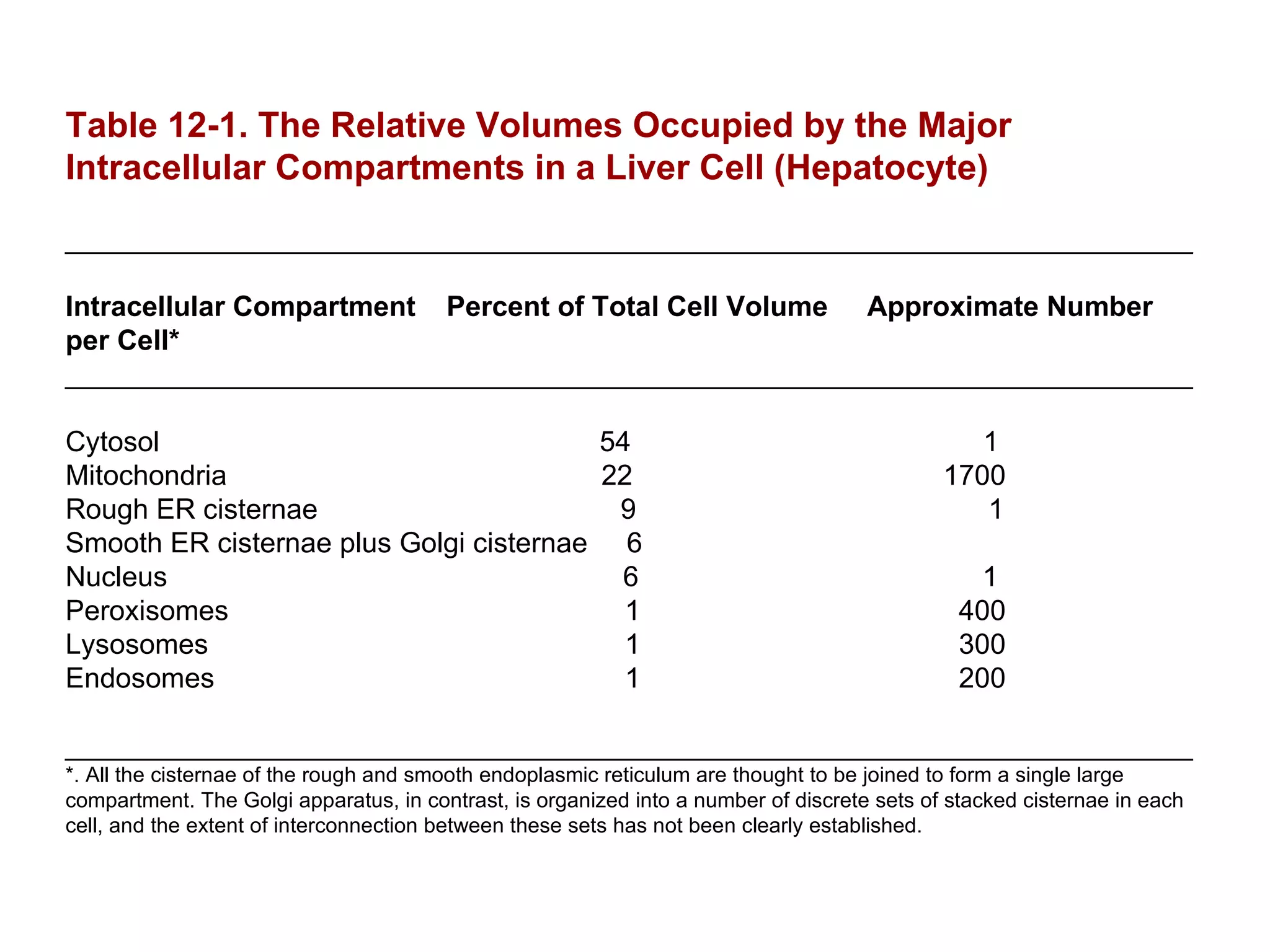
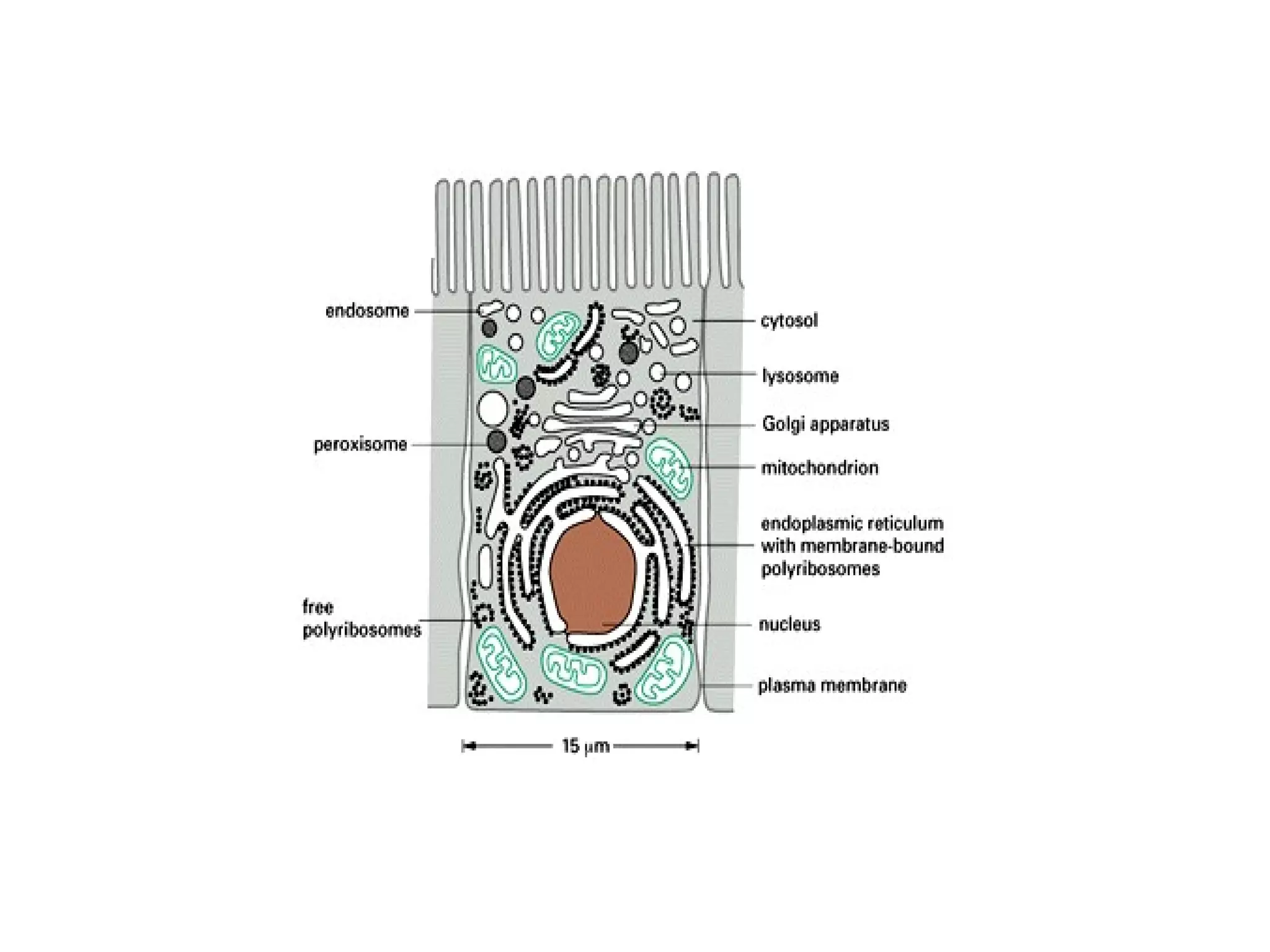
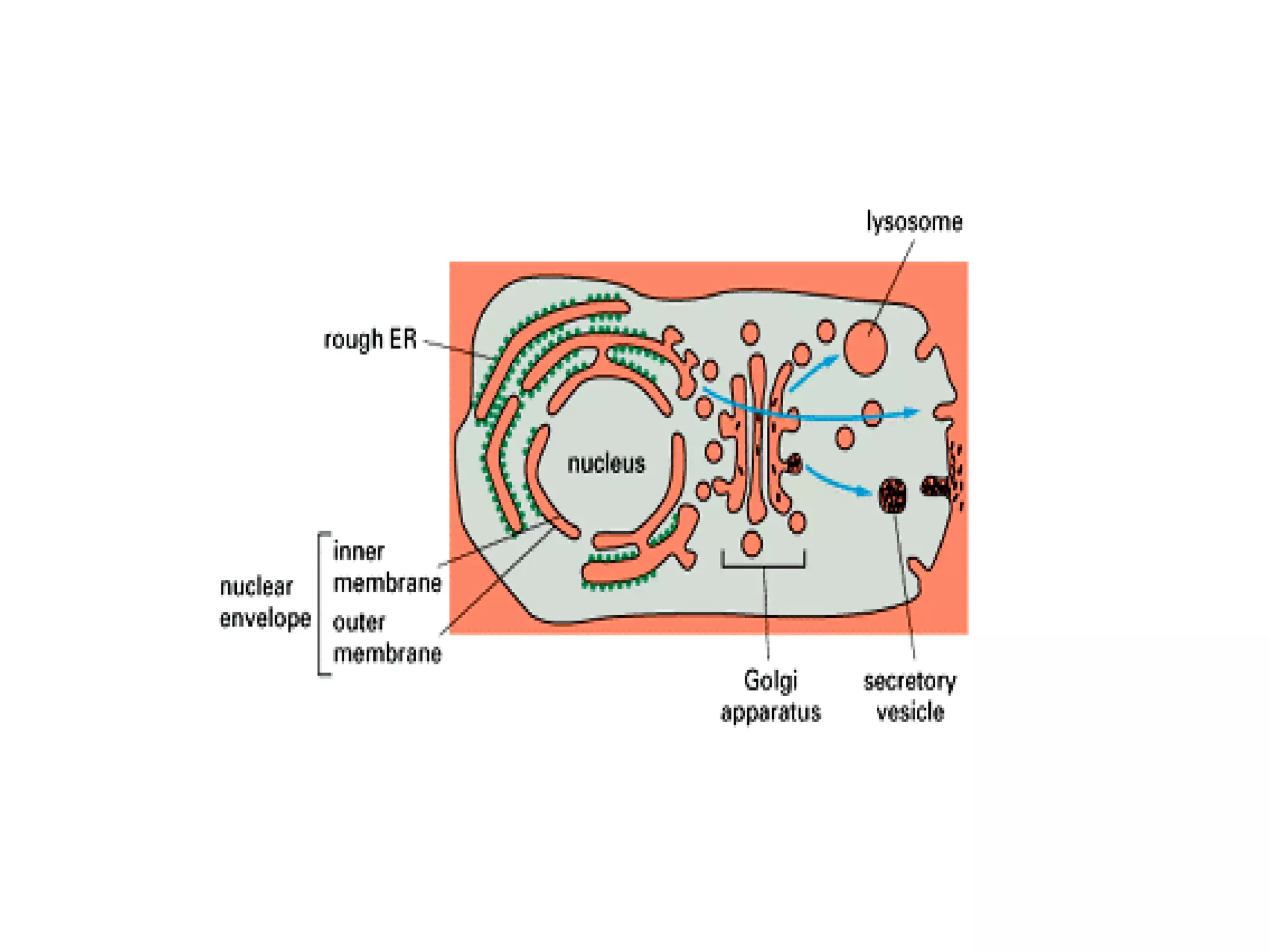
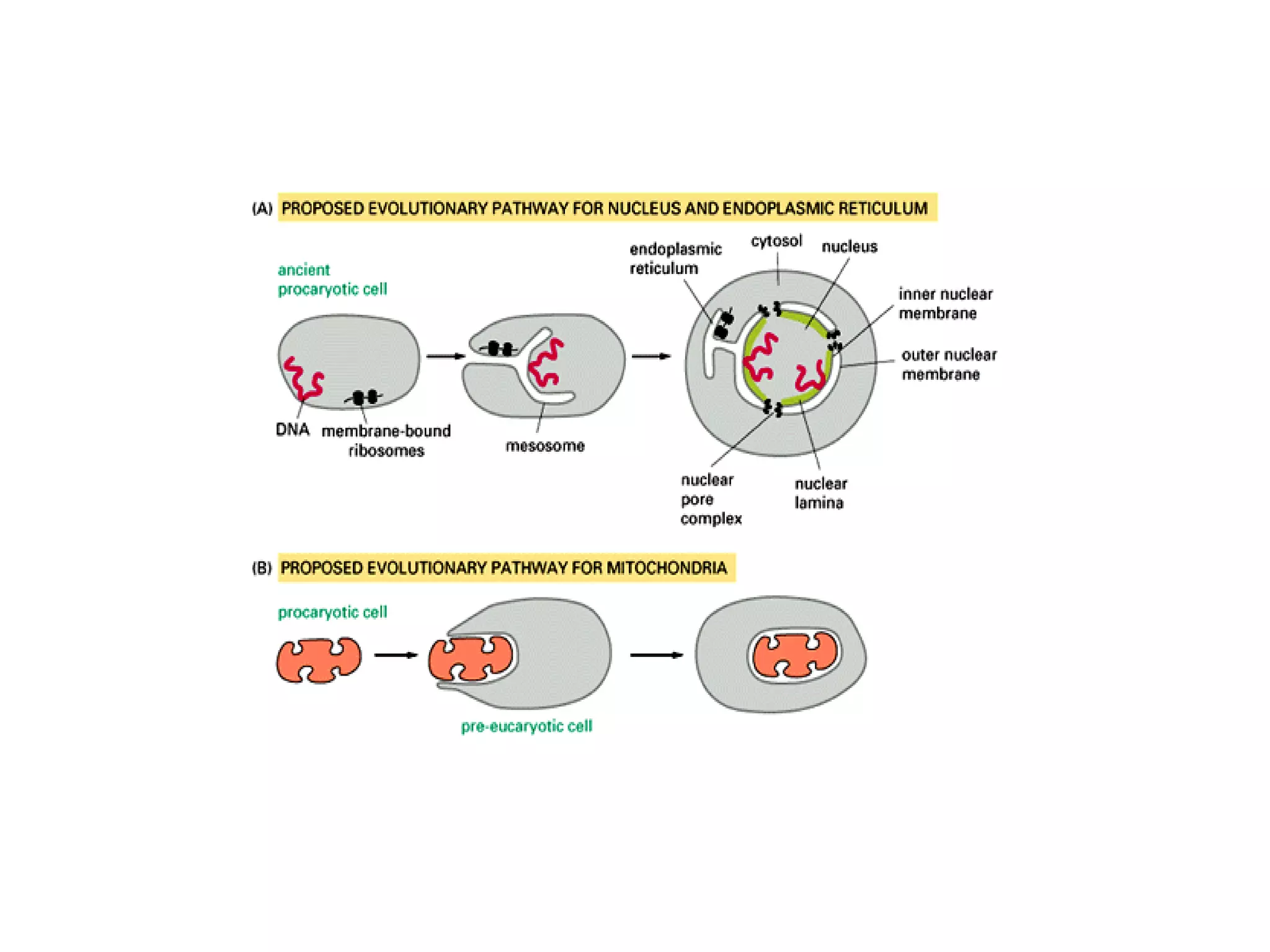
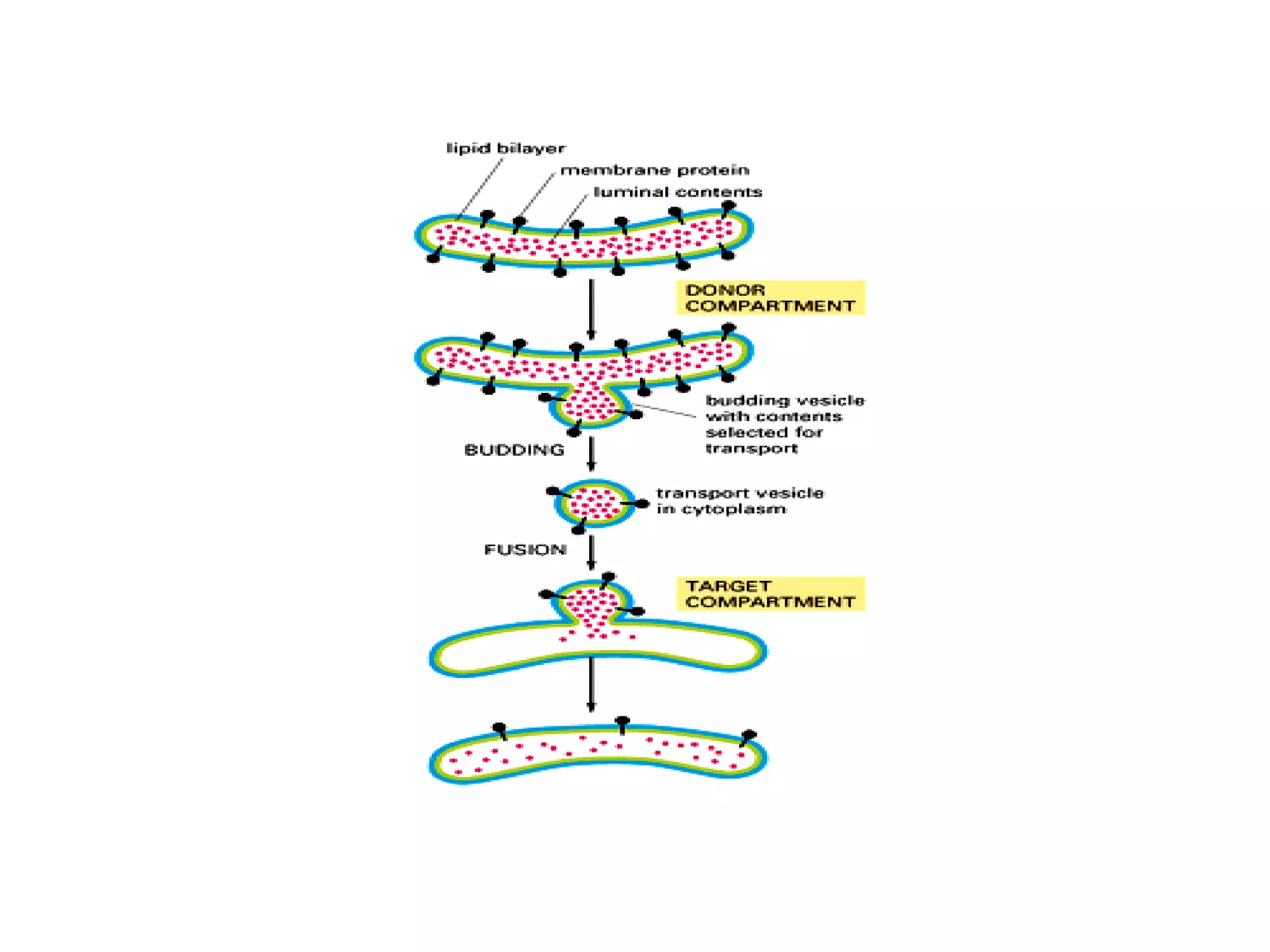
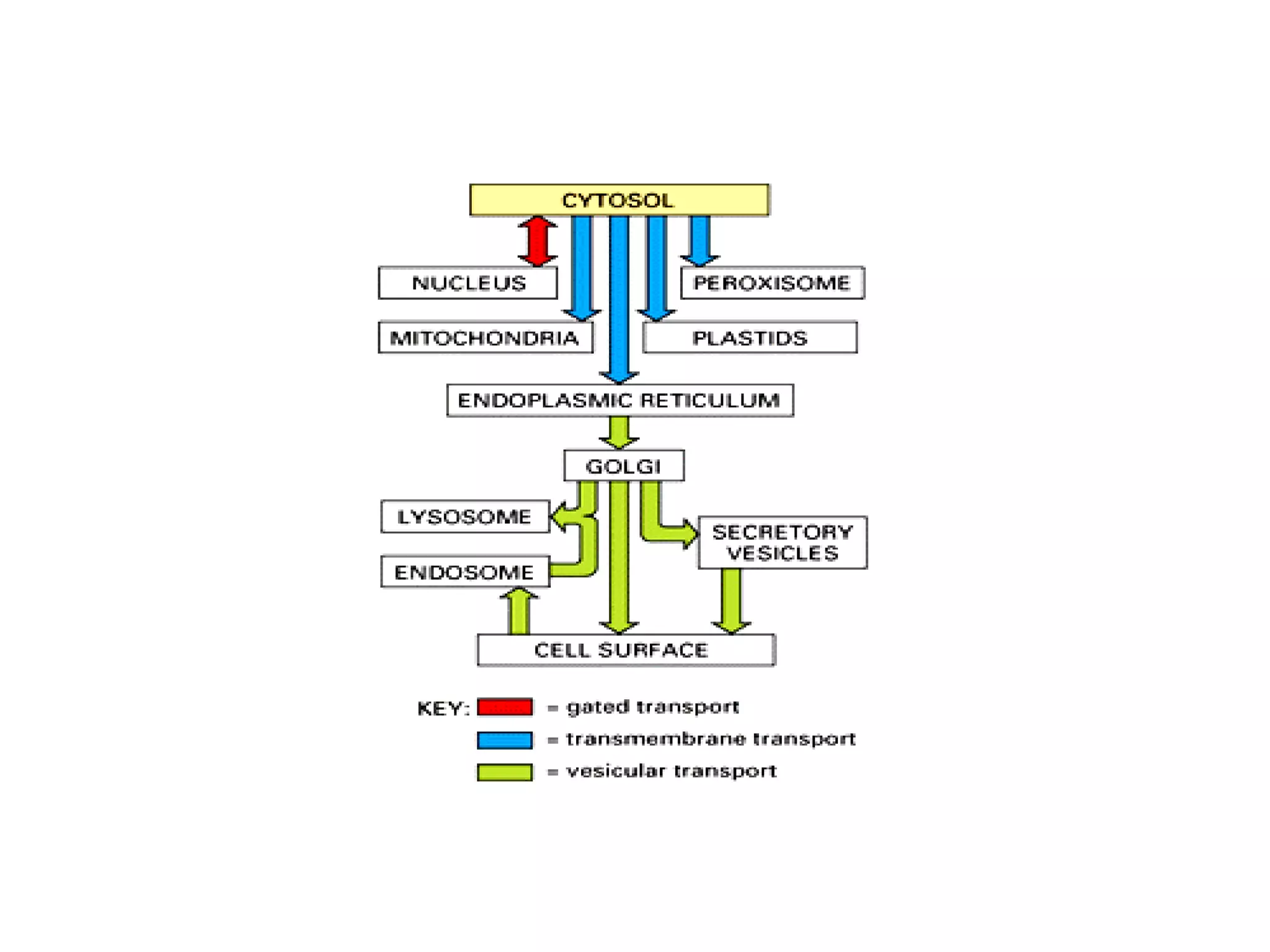
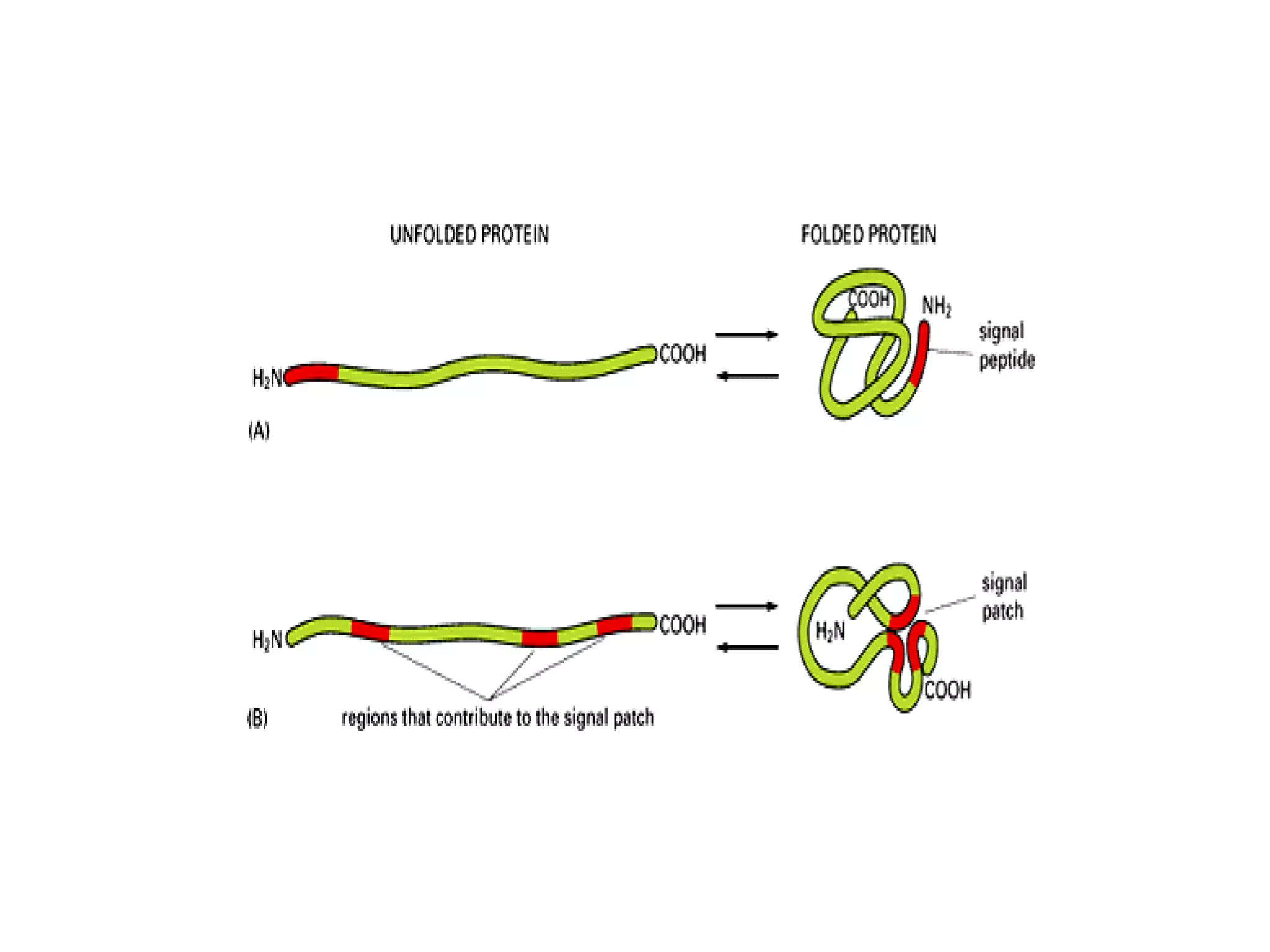
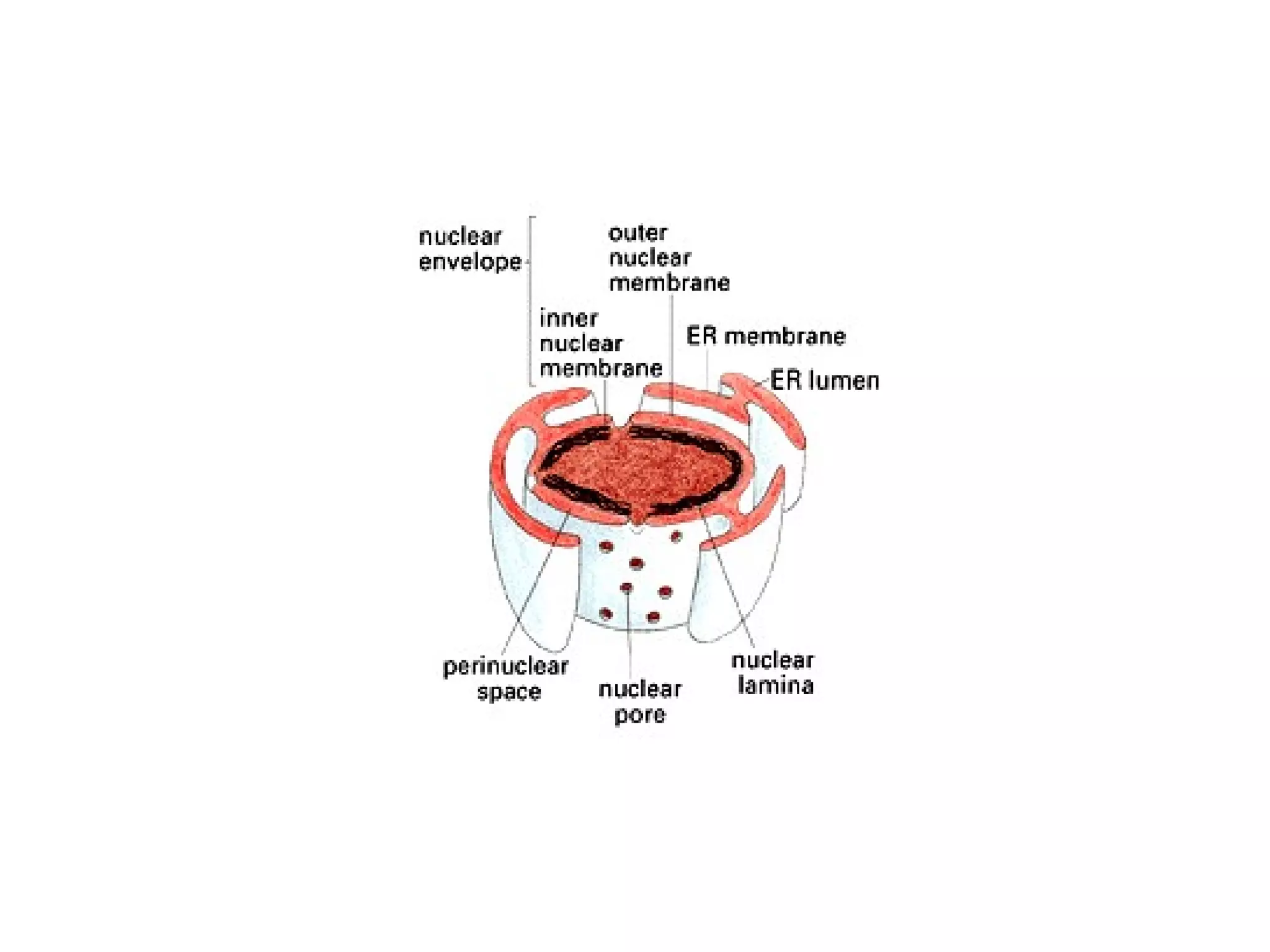
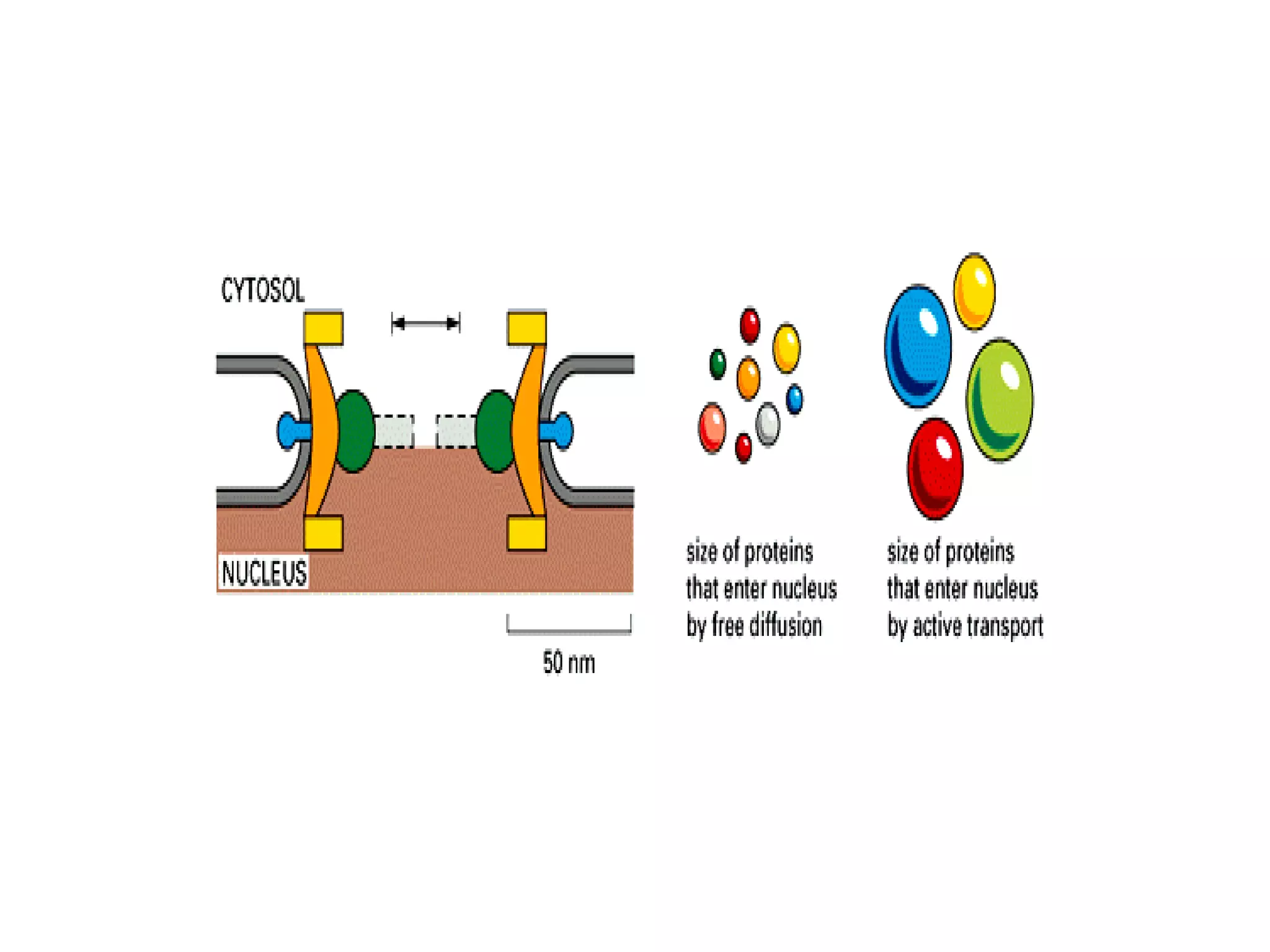
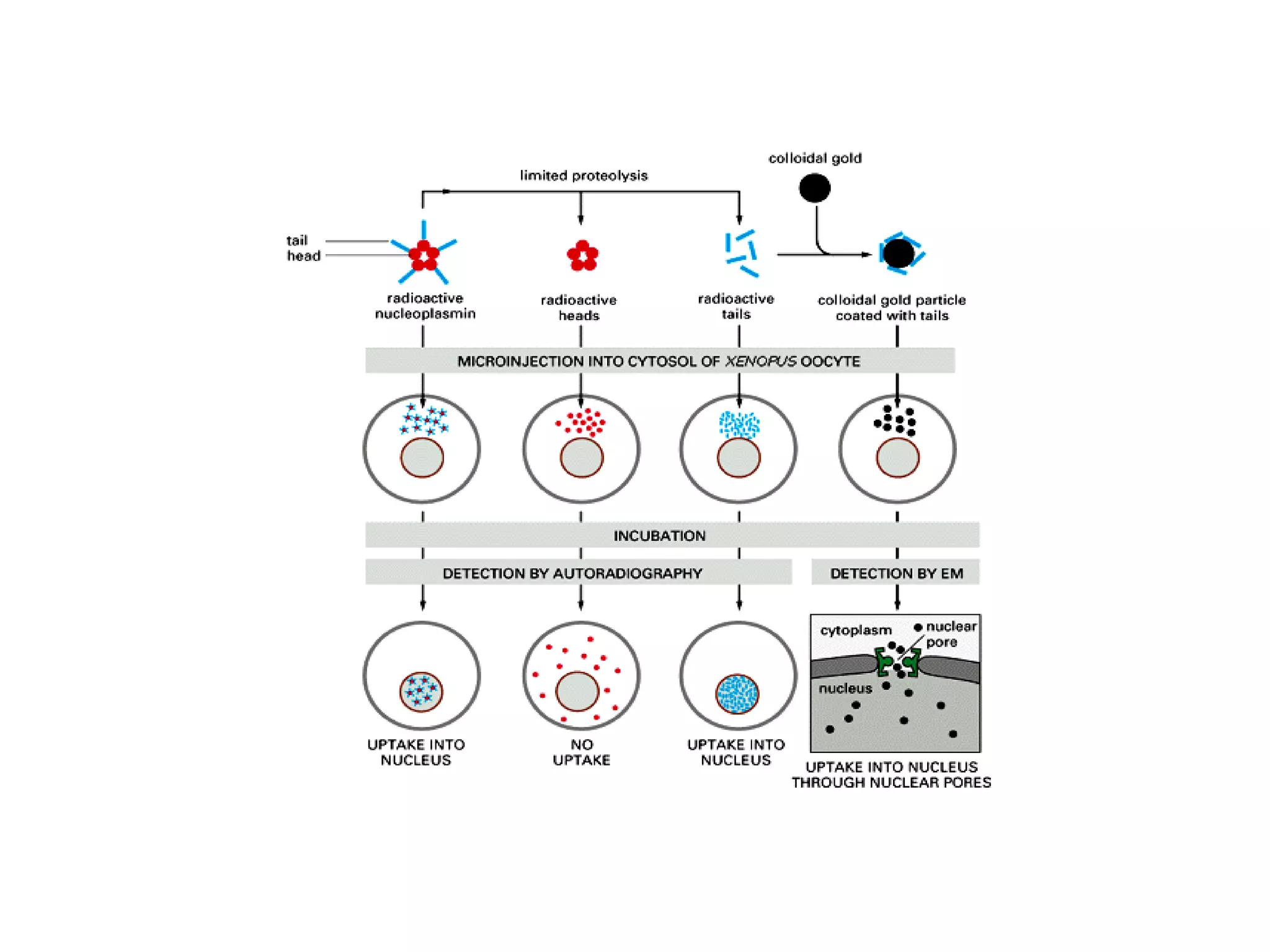
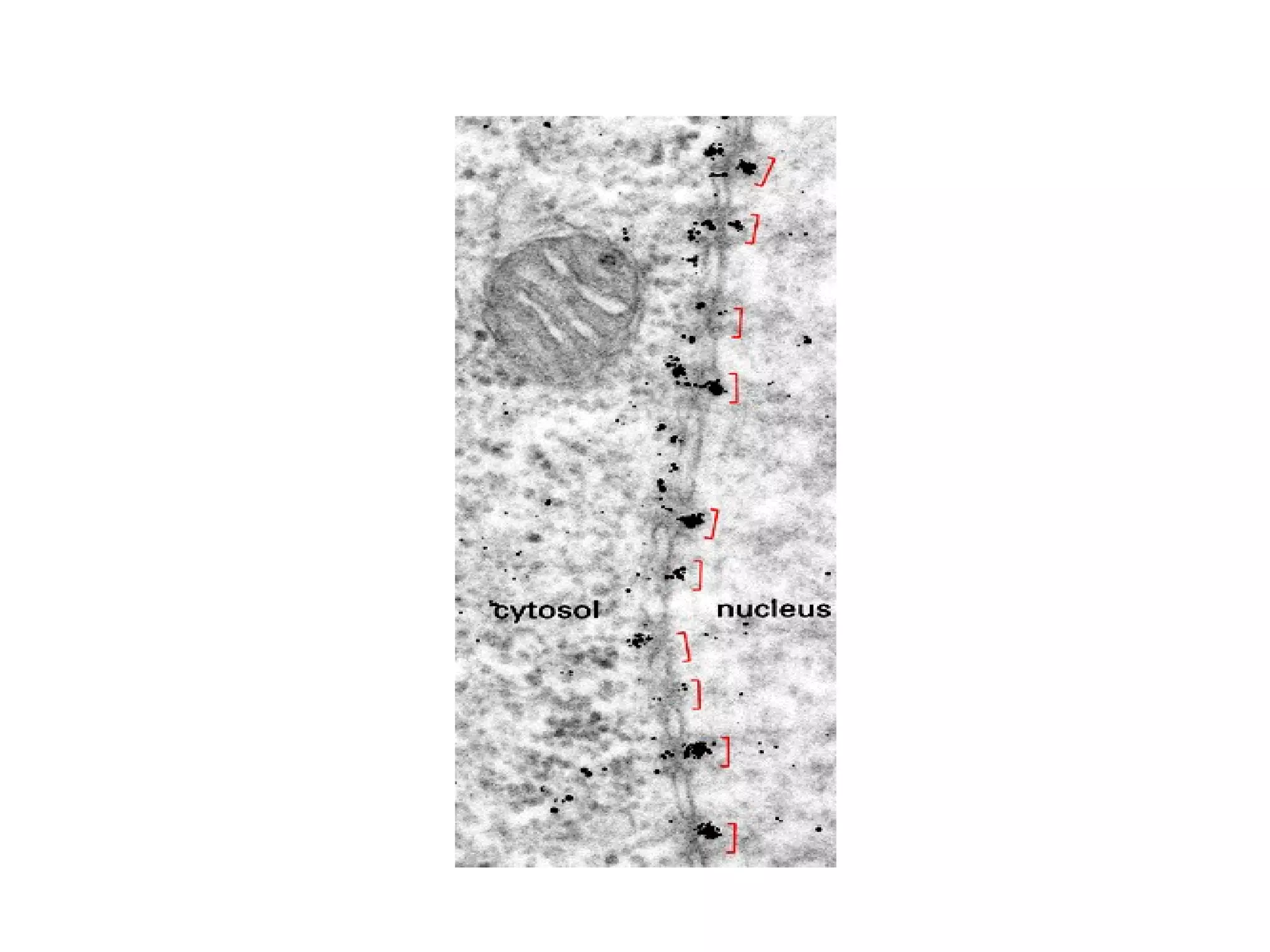
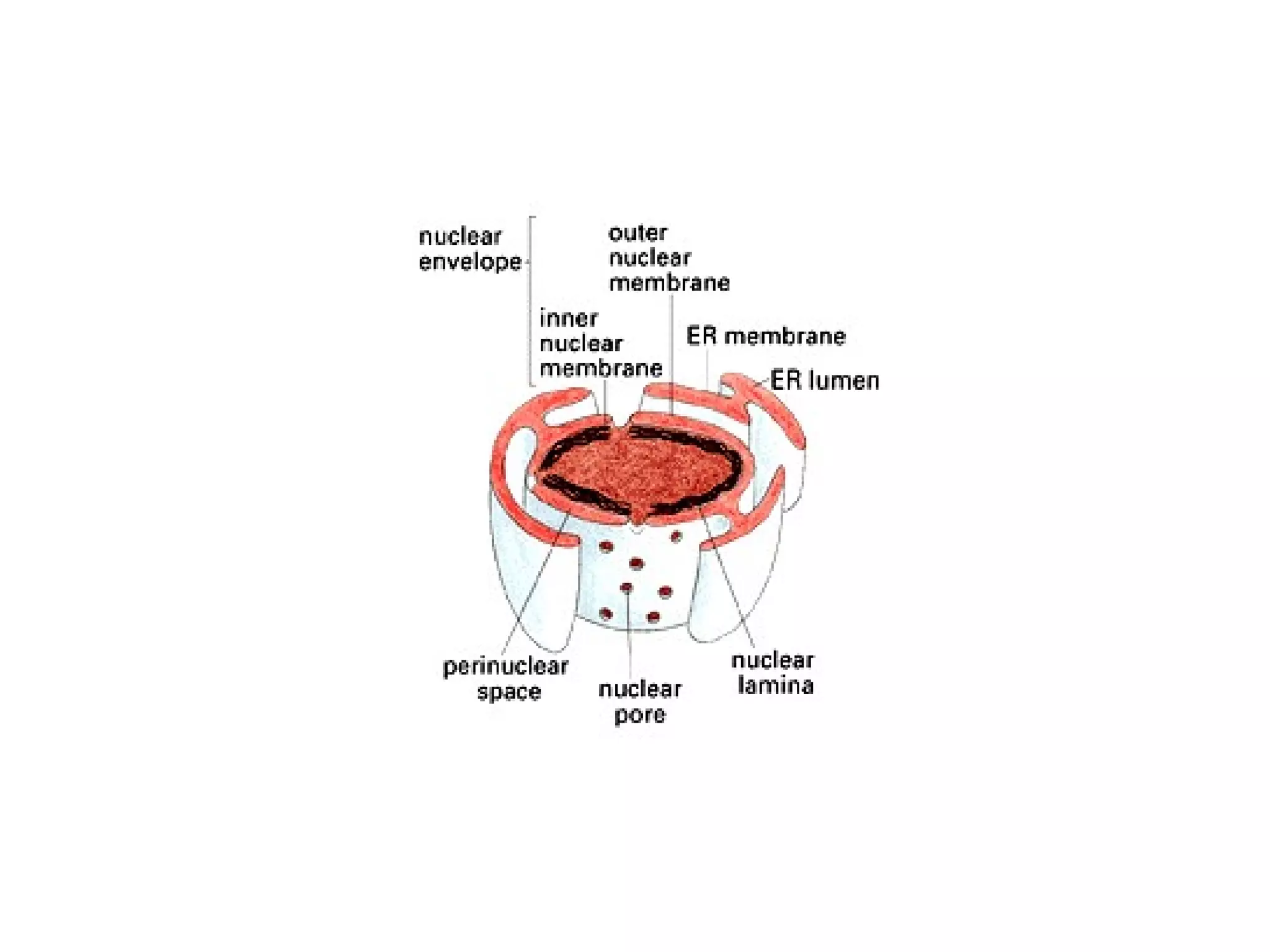
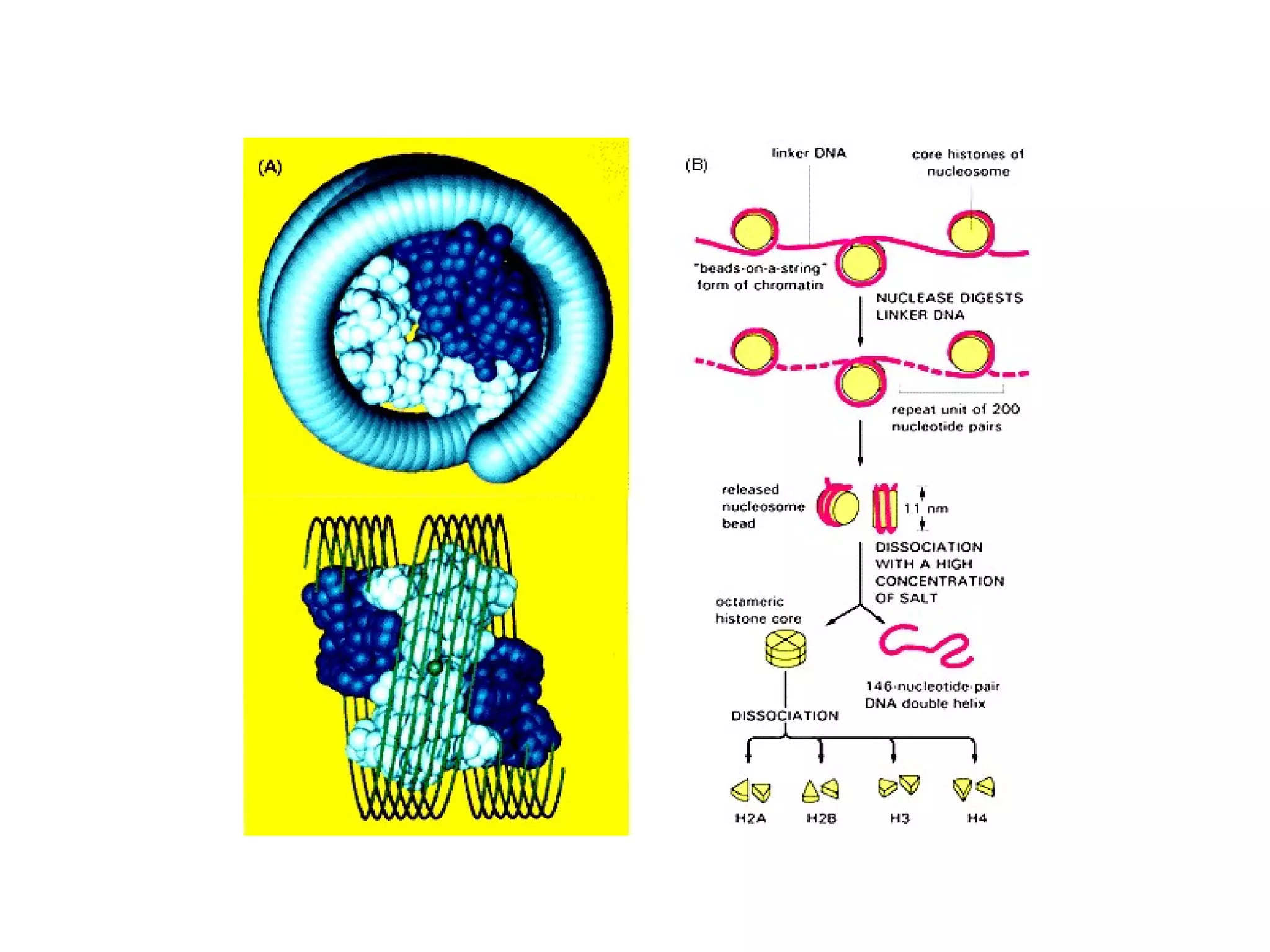
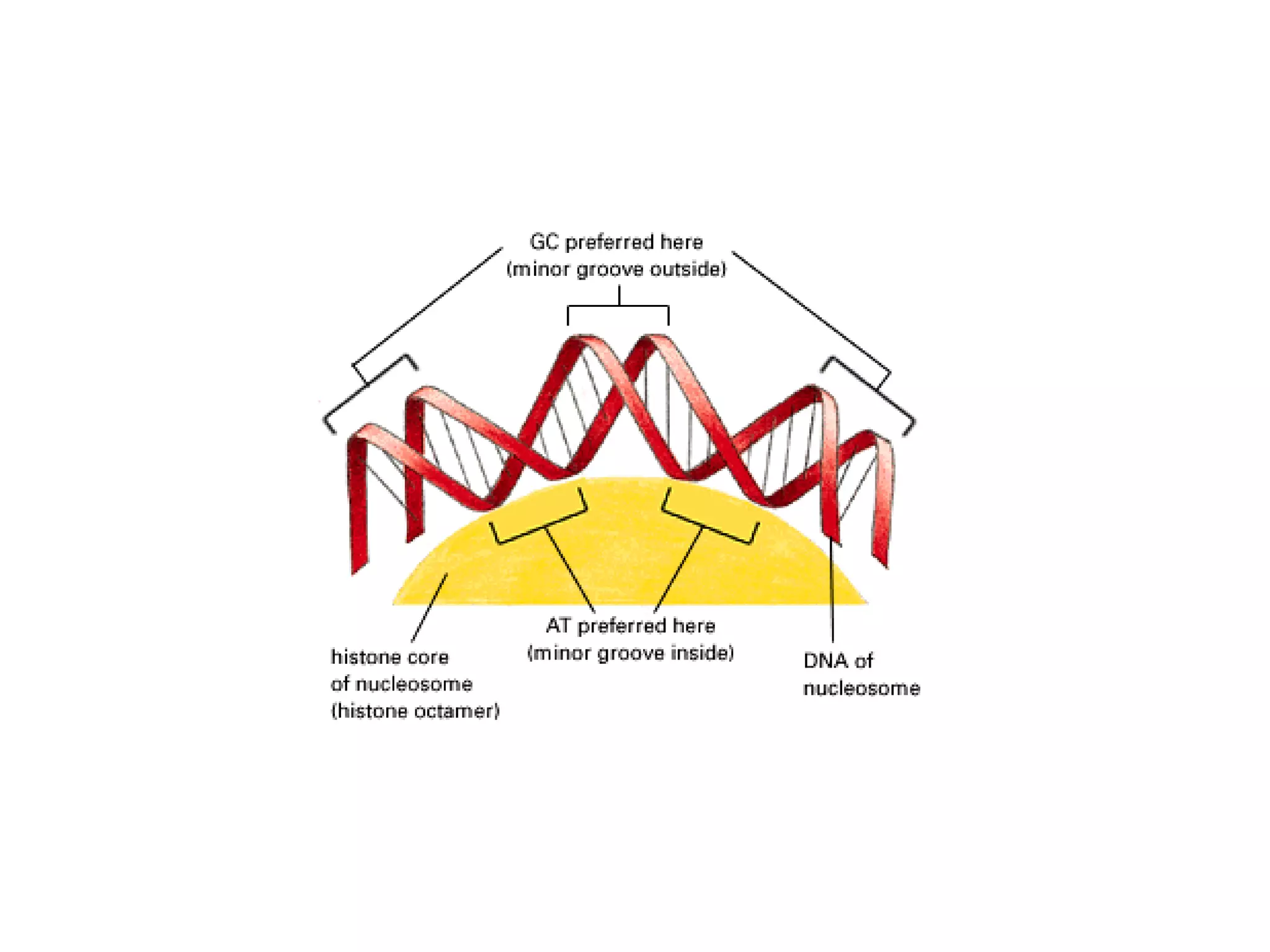
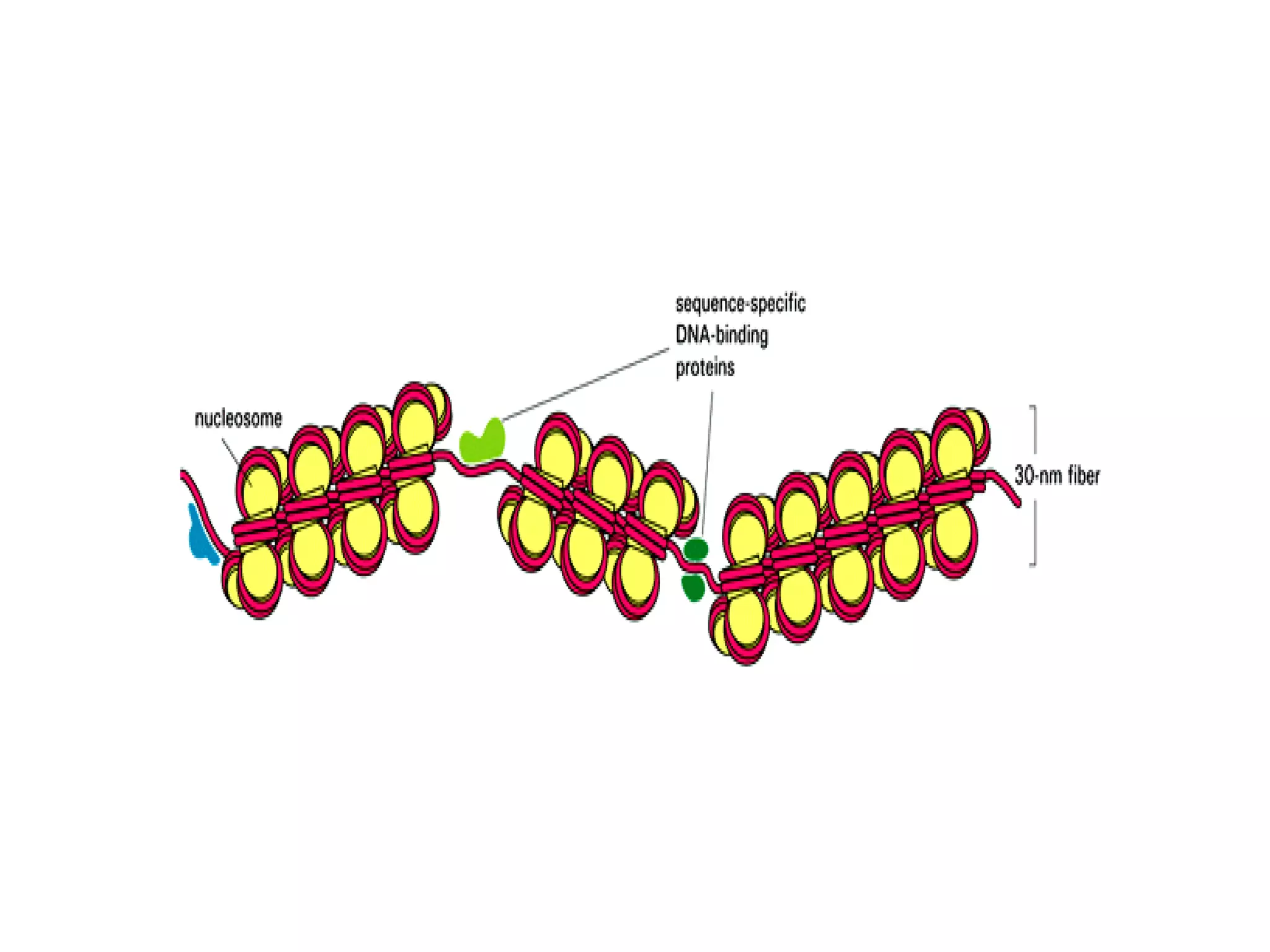
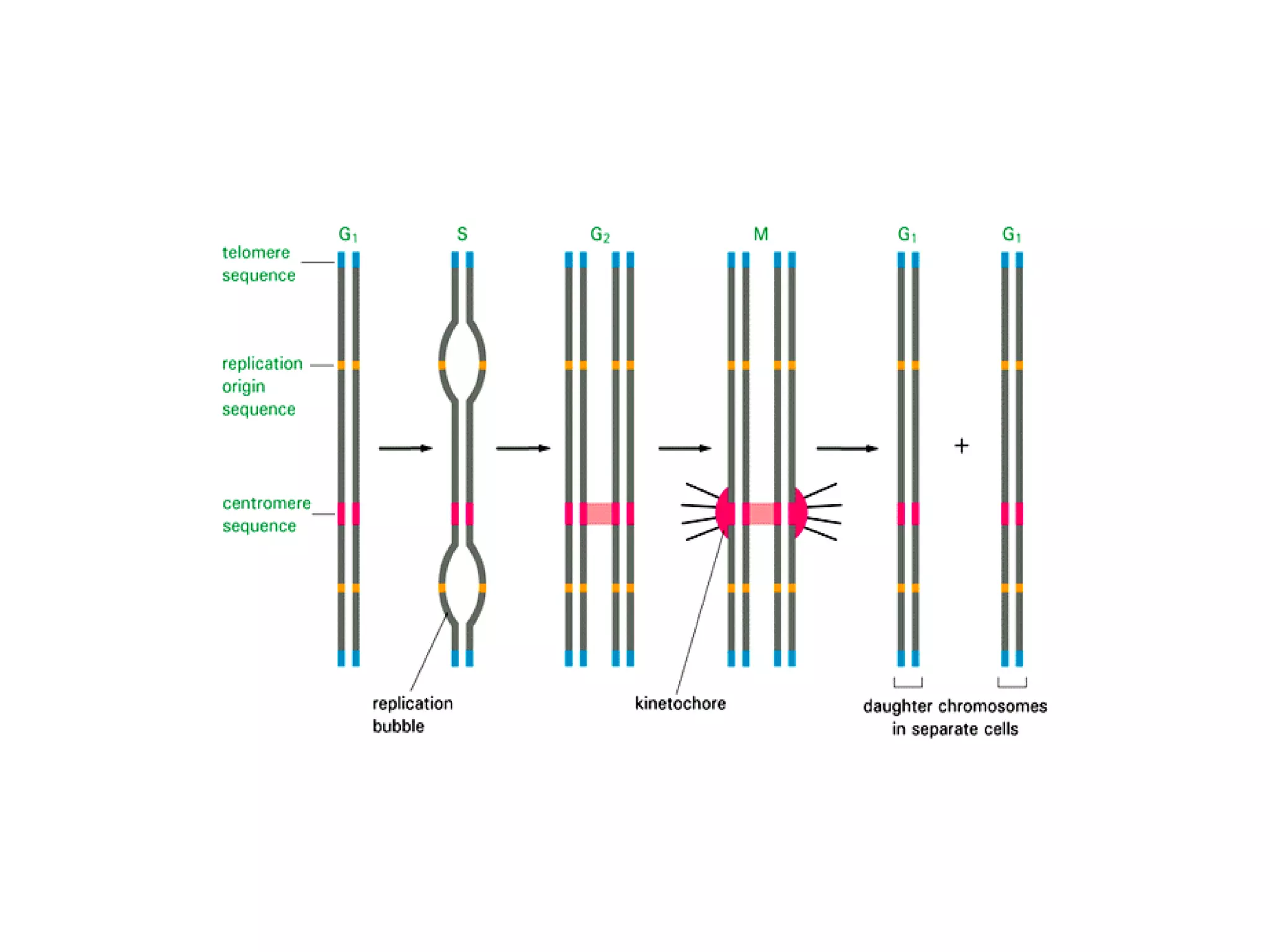
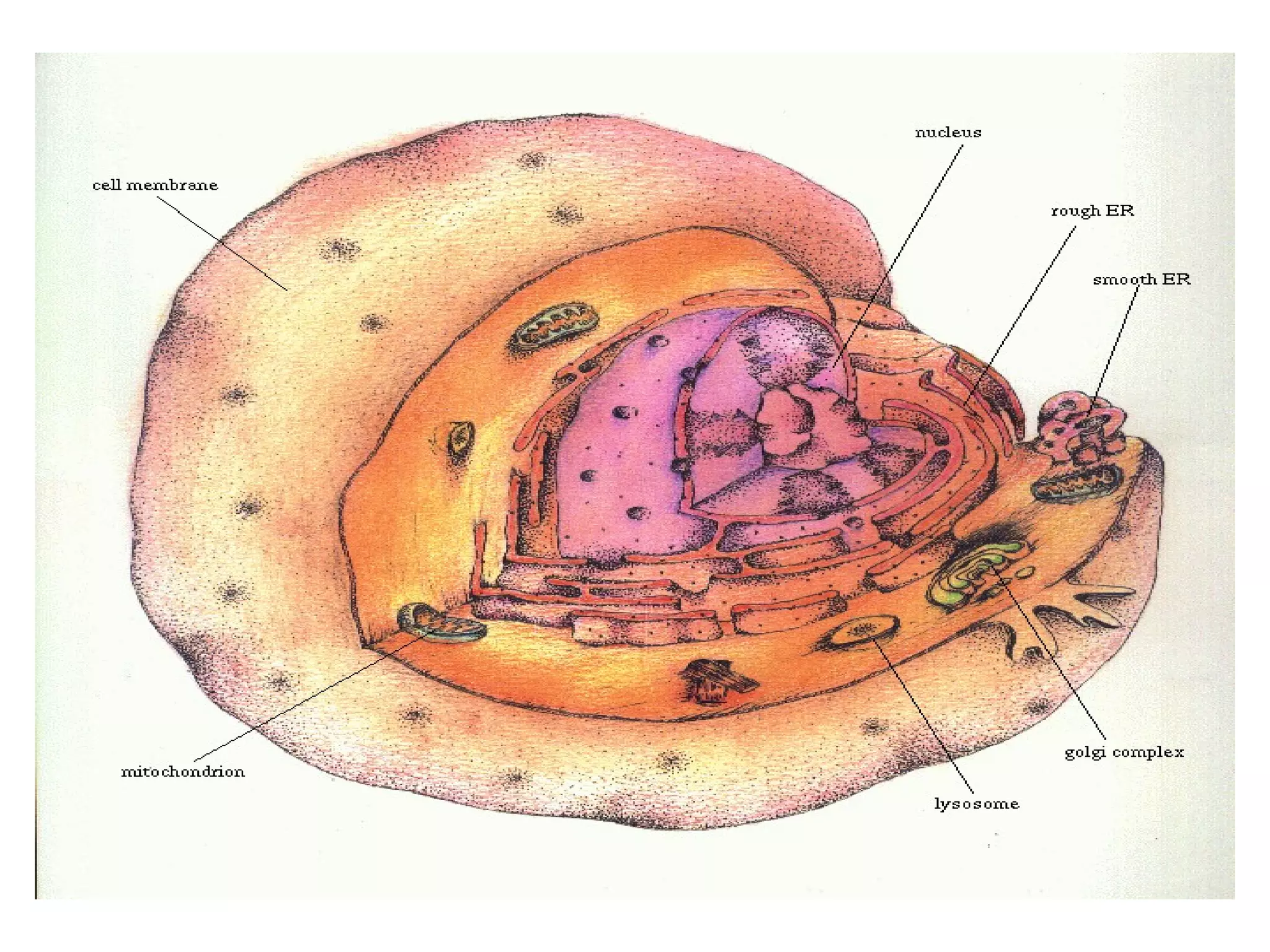
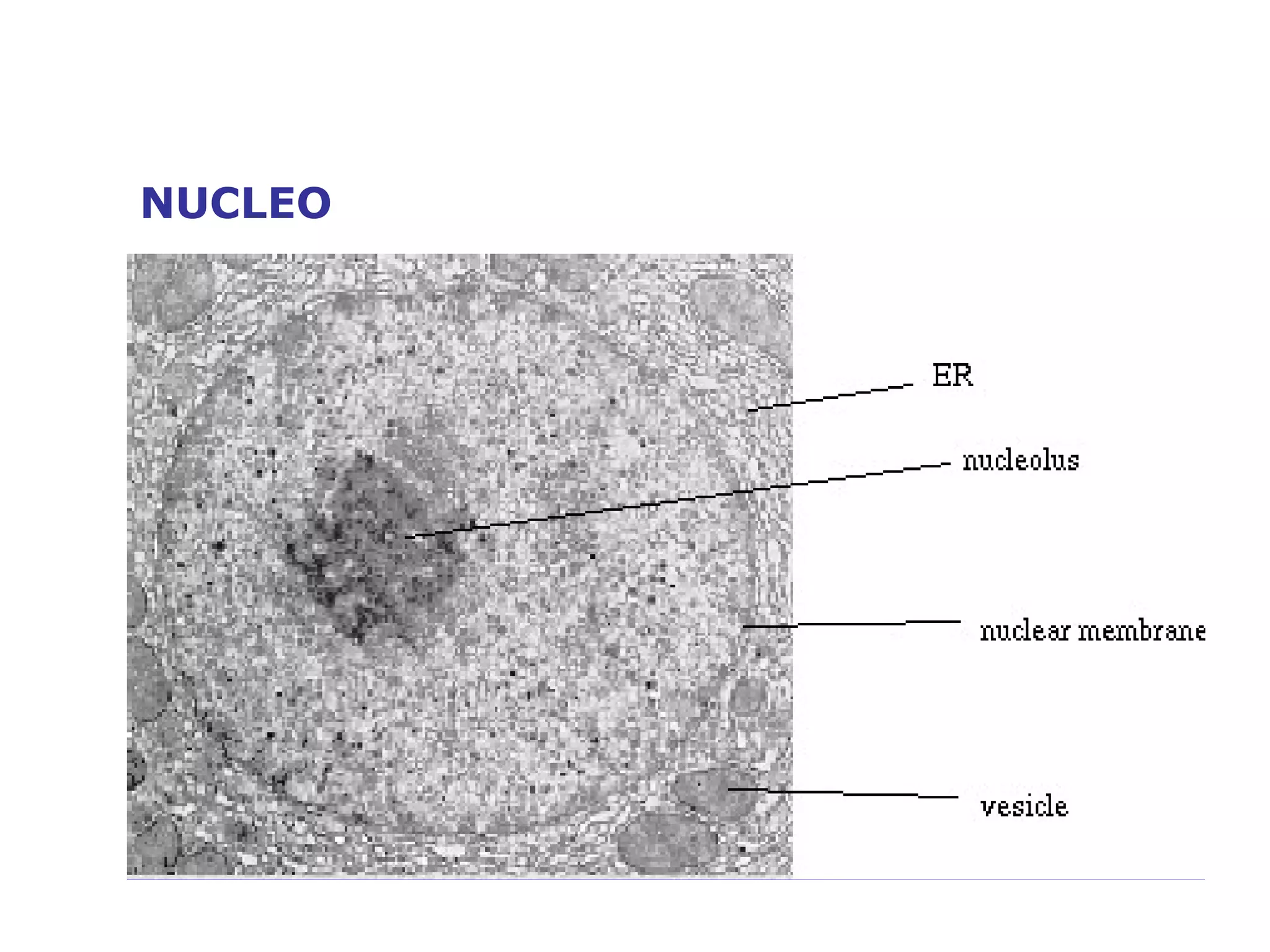
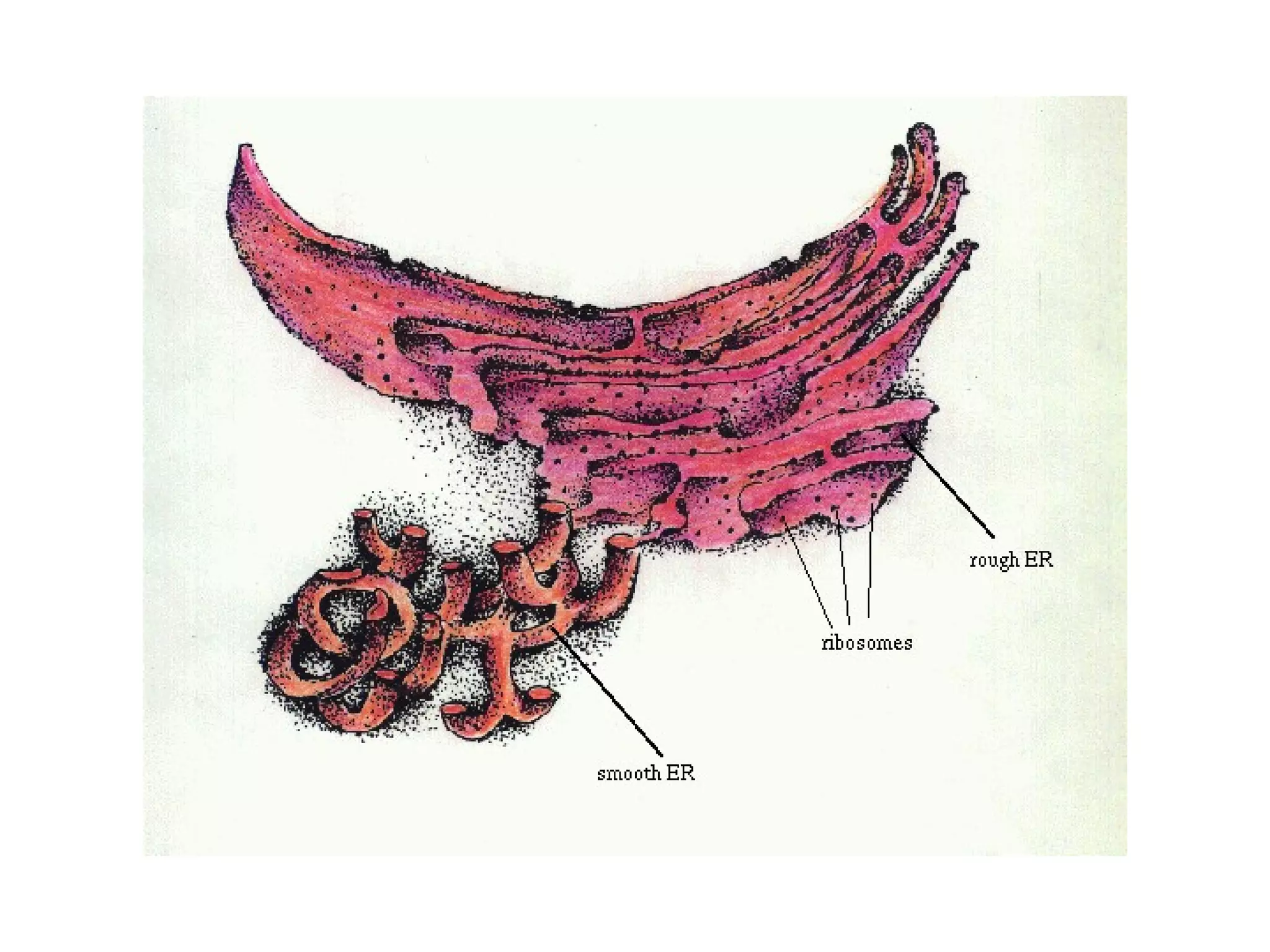
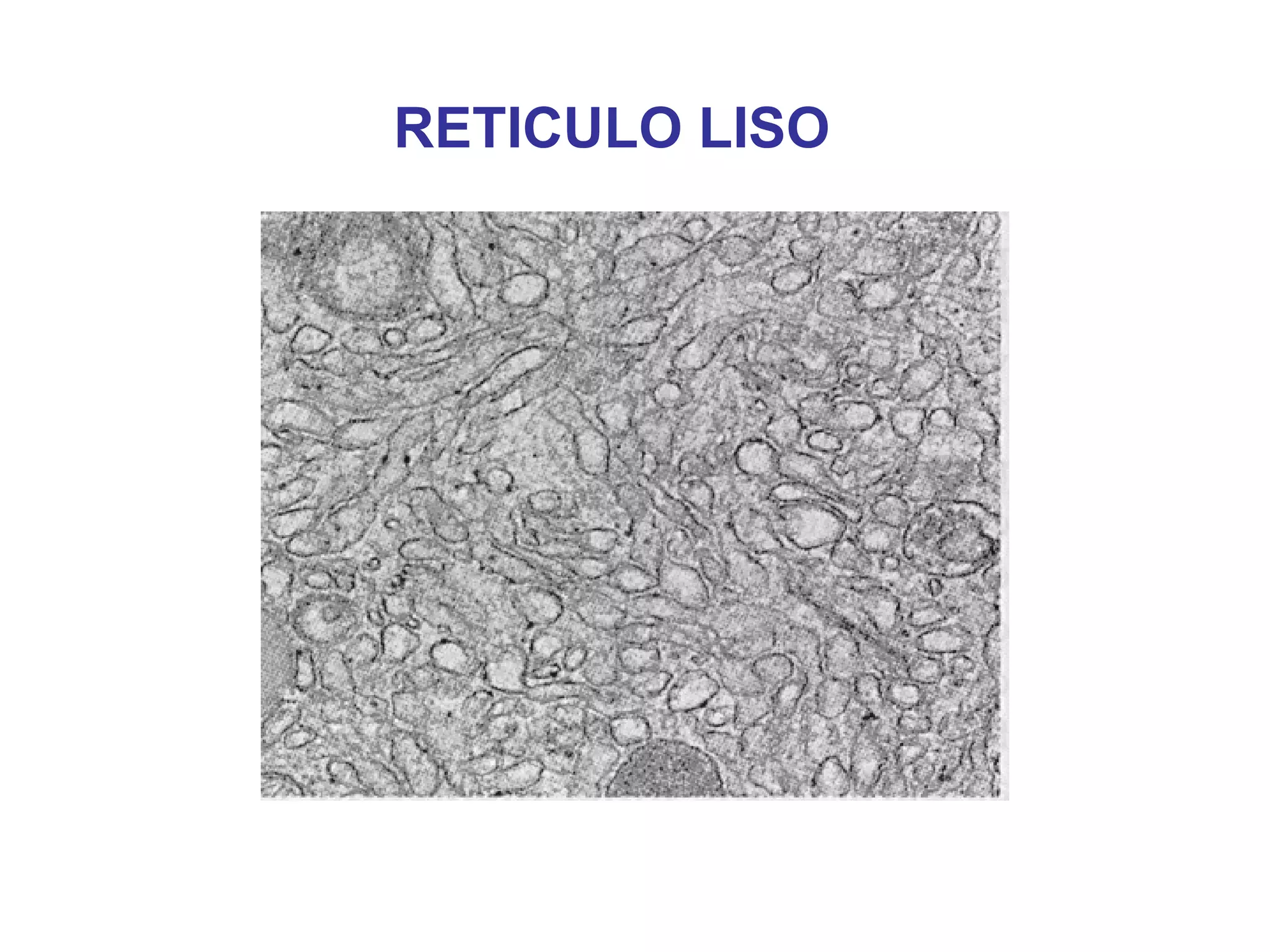
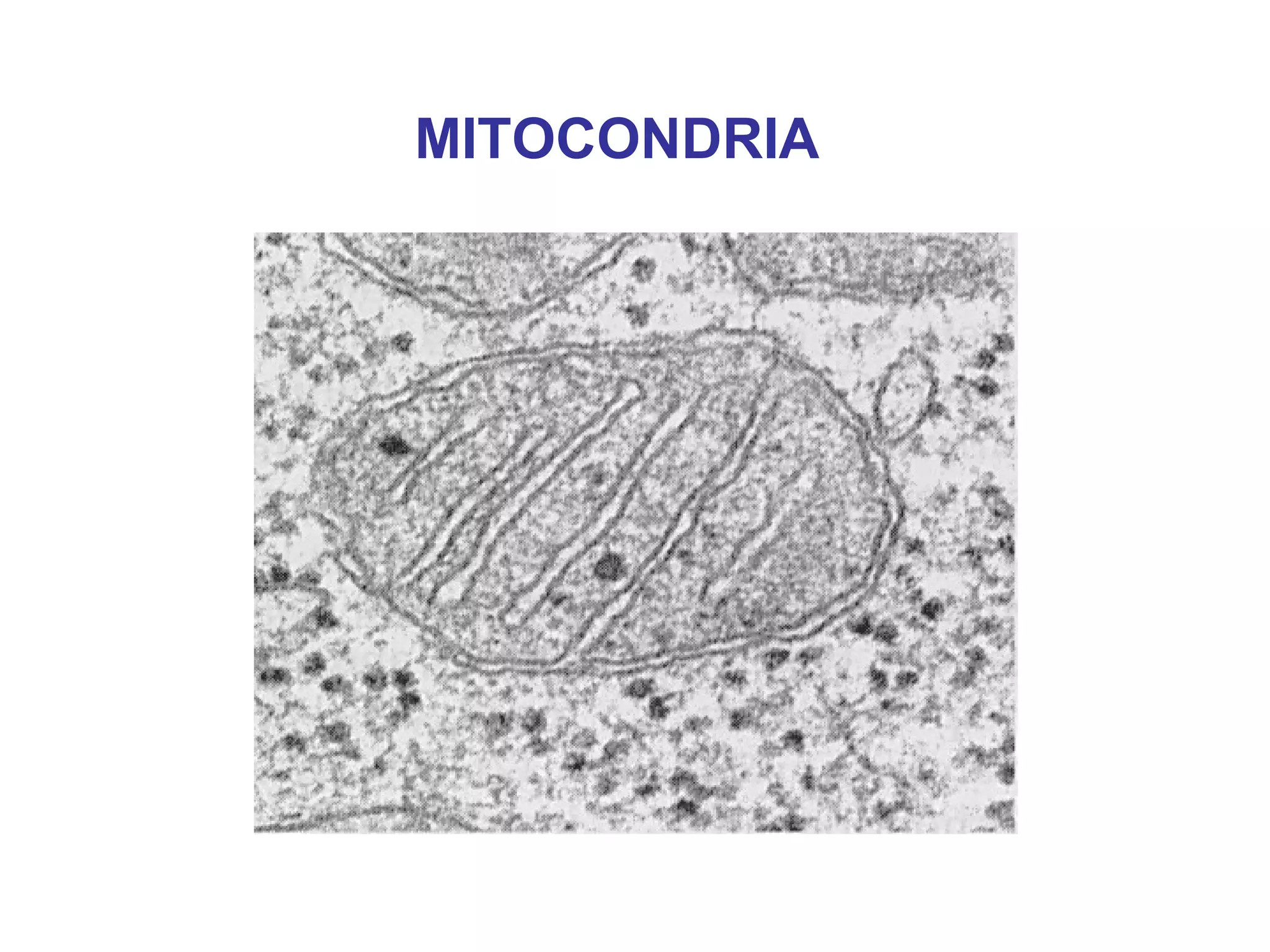
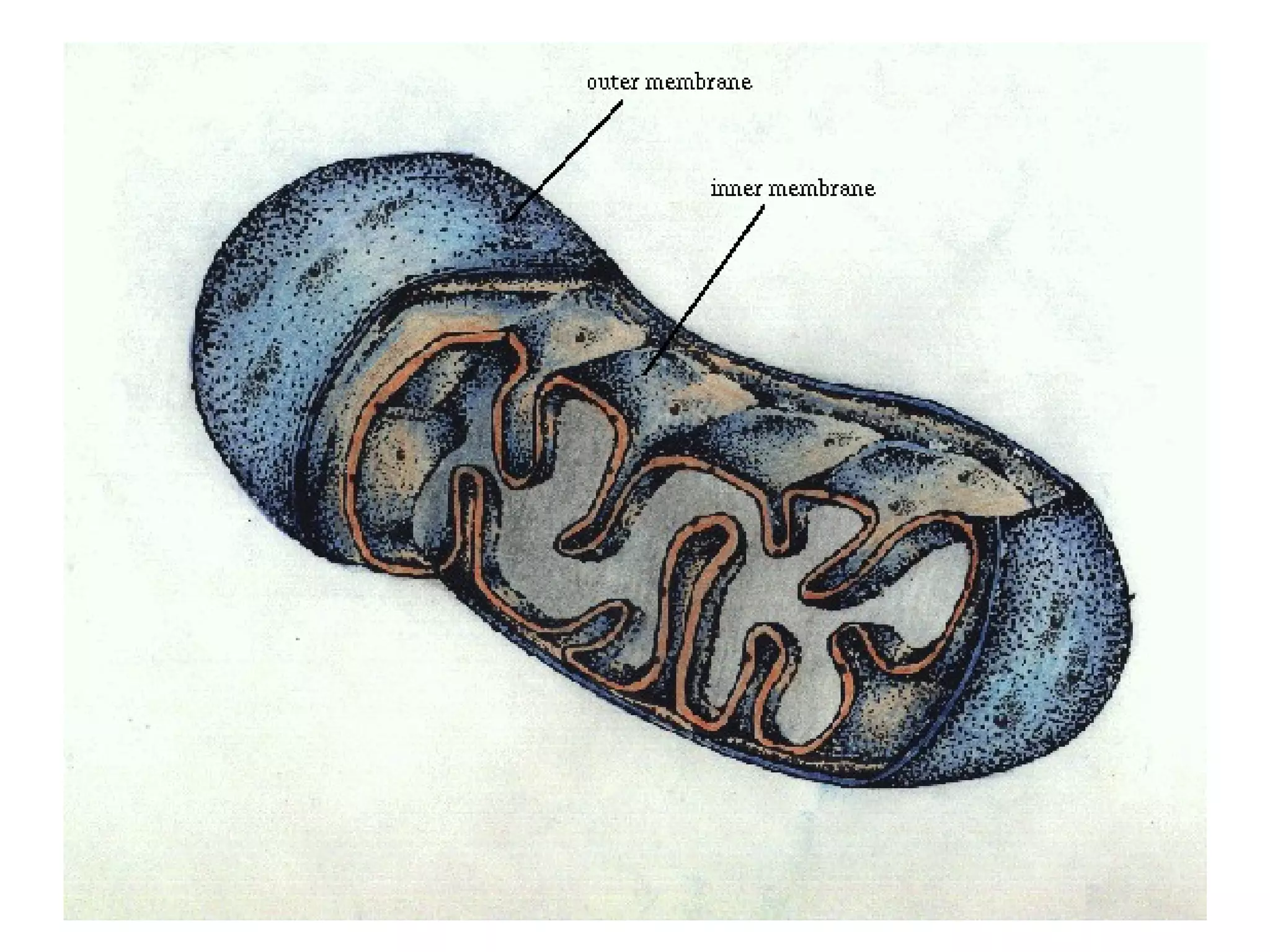
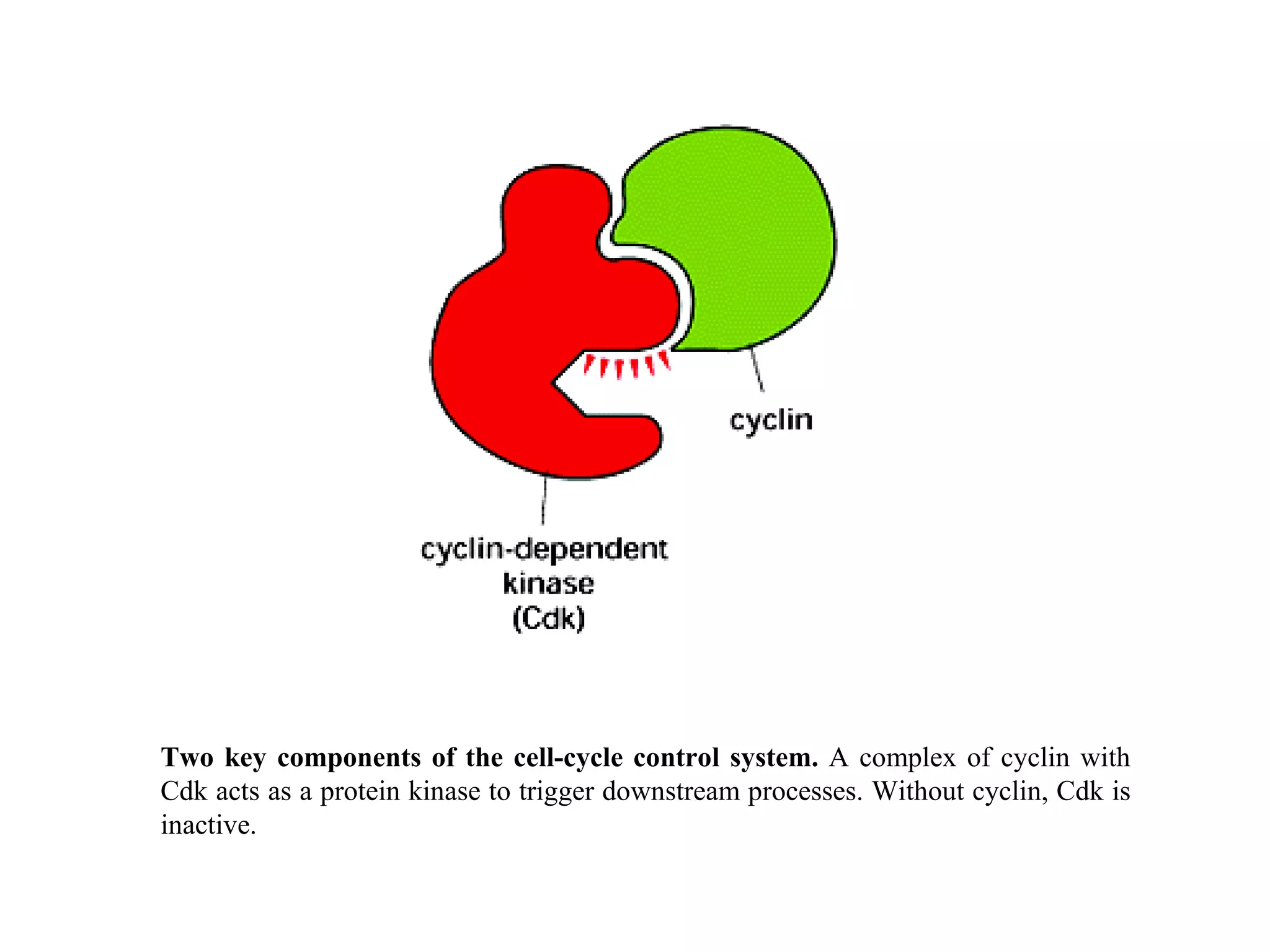
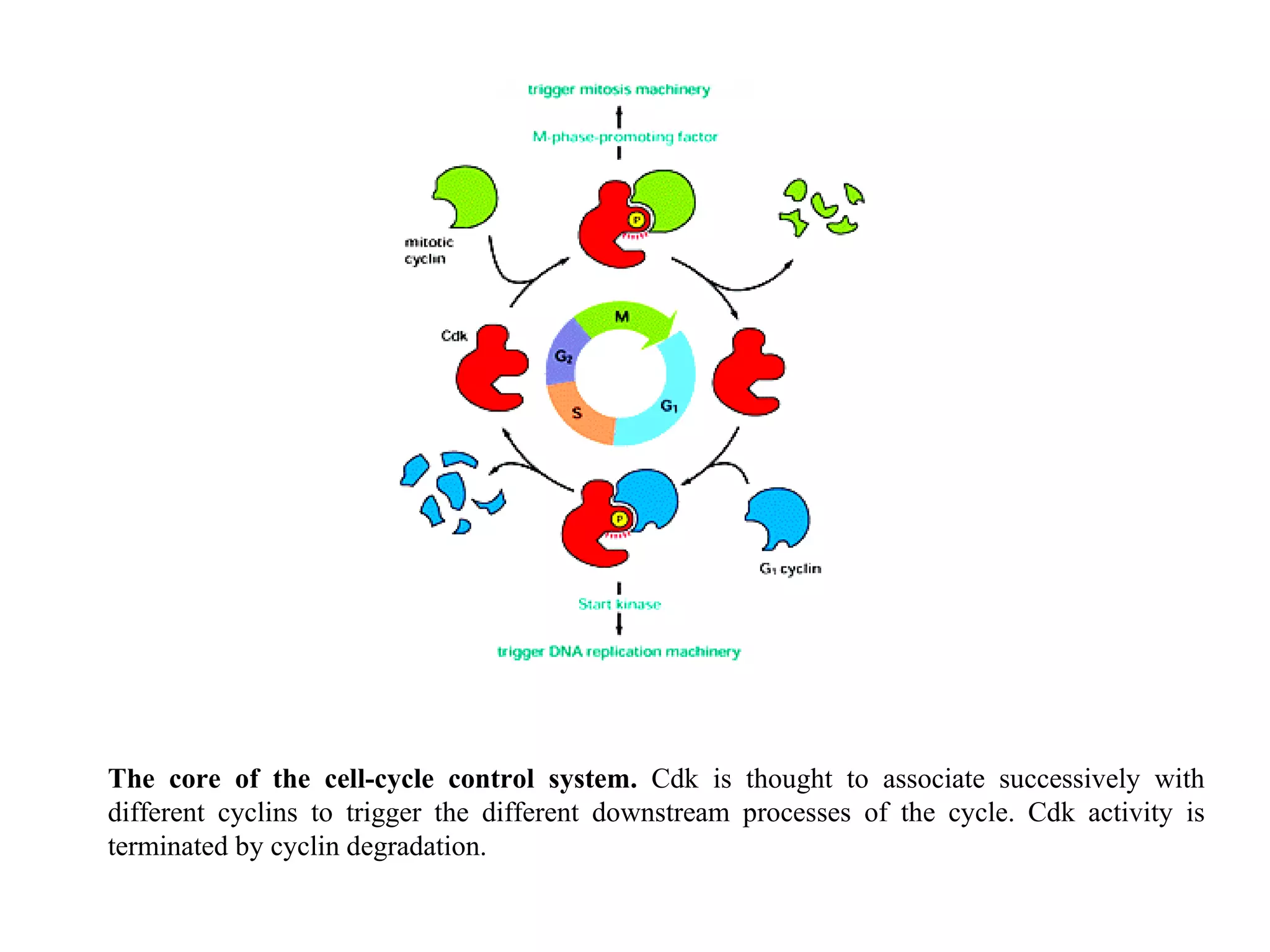
The document describes the intracellular organization of a liver cell (hepatocyte). It includes a table showing the relative volumes occupied by major intracellular compartments, including the cytosol (54% of cell volume), mitochondria (22%), rough endoplasmic reticulum (9%), and nucleus (6%). It notes that the endoplasmic reticulum forms a single large compartment, while the Golgi apparatus is organized into discrete stacked cisternae.






















































































