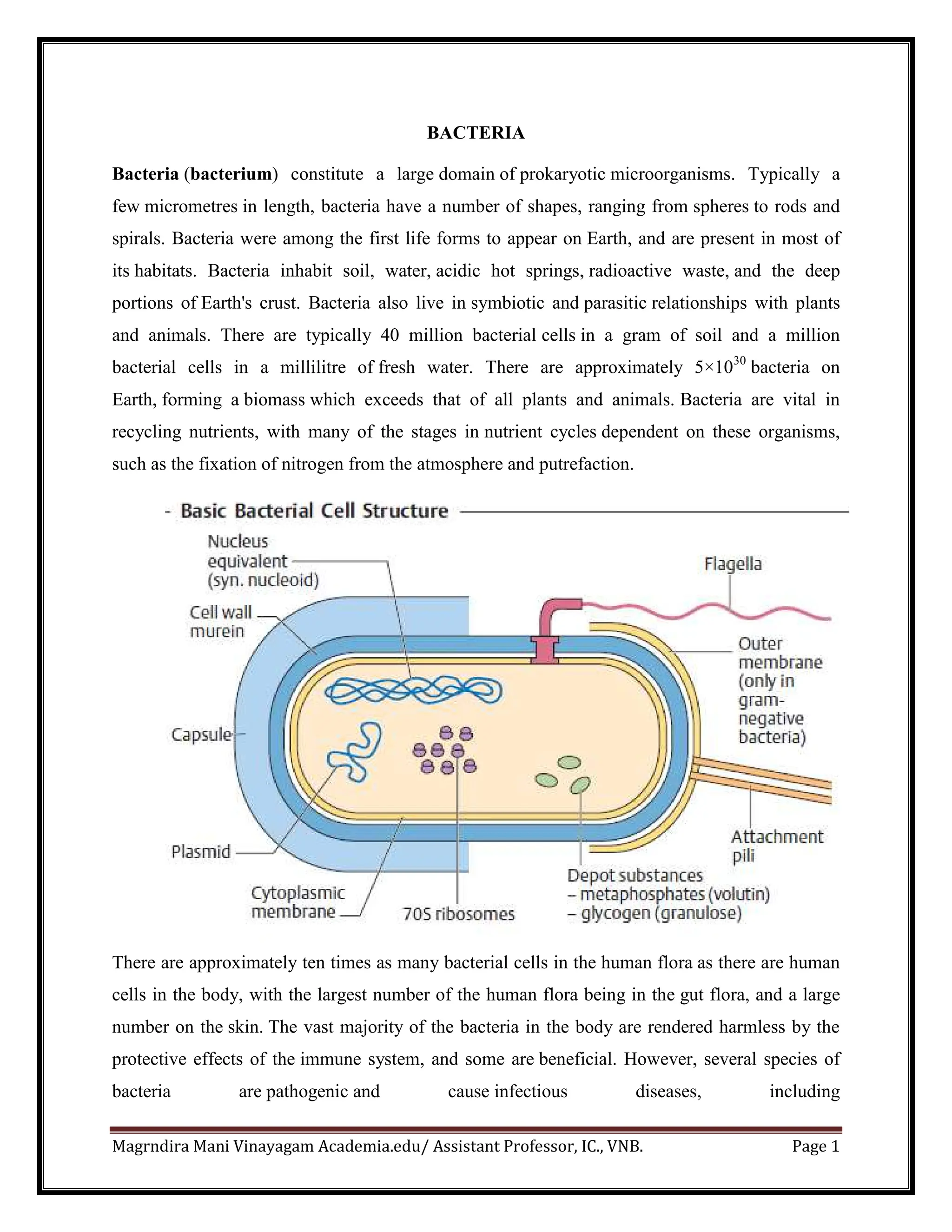
Bacteria are prokaryotic microorganisms with diverse shapes and play essential roles in nutrient recycling, human health, and industry. They can form symbiotic or pathogenic relationships and have significant implications in medicine due to antibiotic resistance and diseases they cause. Their cellular structure includes a unique peptidoglycan cell wall, various internal components, and diverse adaptations for survival.































![sturcture of bacteria lecture 3[1].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/sturctureofbacterialecture31-240128072427-20b3d95c-thumbnail.jpg?width=640&height=640&fit=bounds)












































