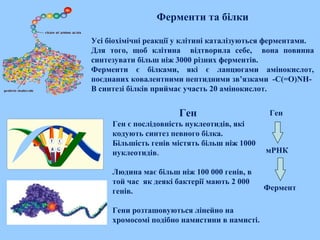
Ферменти та білки
Усібіохімічні реакції у клітині каталізуються ферментами.
Для того, щоб клітина відтворила себе, вона повинна
синтезувати більш ніж 3000 різних ферментів.
Ферменти є білками, які є ланцюгами амінокислот,
поєднаних ковалентними пептидними зв’язками -C(=O)NH-
В синтезі білків приймає участь 20 амінокислот.
Ген
Ген є послідовність нуклеотидів, які
кодують синтез певного білка.
Більшість генів містять більш ніж 1000
нуклеотидів.
Людина має більш ніж 100 000 генів, в
той час як деякі бактерії мають 2 000
генів.
Гени розташовуються лінейно на
хромосомі подібно намистини в намисті.
Ген
мРНК
Фермент
3.
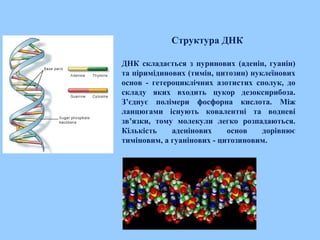
Структура ДНК
ДНК складаєтьсяз пуринових (аденін, гуанін)
та піримідинових (тимін, цитозин) нуклеїнових
основ - гетероциклічних азотистих сполук, до
складу яких входить цукор дезоксирибоза.
З’єднує полімери фосфорна кислота. Між
ланцюгами існують ковалентні та водневі
зв’язки, тому молекули легко розпадаються.
Кількість аденінових основ дорівнює
тиміновим, а гуанінових - цитозиновим.
4.
Довжина прокаріотної ДНК
Якщовзяти ДНК із однієї клітини людини, то її довжина буде приблизно
дорівнюватися 1м.
Довжина спрямленої ДНК від Escherichia coli дорівнює приблизно 1000 µm,
хоч довжина самої бактеріальної клітини складає 2 µм.
ДНК із Escherichia coli має 4 мільйонів пар нуклеотидних основ.
ДНК із однієї клітини людини містить біля 3 мільярдів пар основ.
Бактеріальна клітина має одну хромосому, яка є кільцевою молекулою
ДНК.
5.
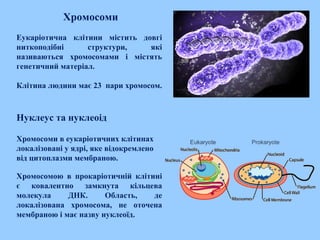
Хромосоми
Еукаріотична клітини міститьдовгі
ниткоподібні структури, які
називаються хромосомами і містять
генетичний матеріал.
Клітина людини має 23 пари хромосом.
Нуклеус та нуклеоід
Хромосоми в еукаріотичних клітинах
локалізовані у ядрі, яке відокремлено
від цитоплазми мембраною.
Хромосомою в прокаріотичній клітині
є ковалентно замкнута кільцева
молекула ДНК. Область, де
локалізована хромосома, не оточена
мембраною і має назву нуклеоїд.
6.
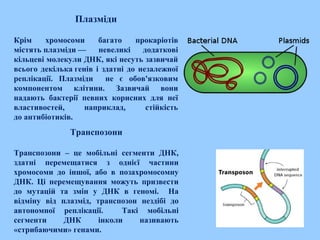
Плазміди
Крім хромосоми багатопрокаріотів
містять плазміди — невеликі додаткові
кільцеві молекули ДНК, які несуть зазвичай
всього декілька генів і здатні до незалежної
реплікації. Плазміди не є обов'язковим
компонентом клітини. Зазвичай вони
надають бактерії певних корисних для неї
властивостей, наприклад, стійкість
до антибіотиків.
Транспозони
Транспозони – це мобільні сегменти ДНК,
здатні перемещатися з однієї частини
хромосоми до іншої, або в позахромосомну
ДНК. Ці перемещування можуть призвести
до мутацій та змін у ДНК в геномі. На
відміну від плазмід, транспозон нездібі до
автономної реплікації. Такі мобільні
сегменти ДНК інколи називають
«стрибаючими» генами.
7.
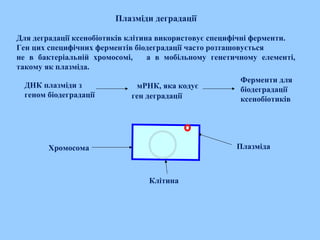
Плазміди деградації
Для деградаціїксенобіотиків клітина використовує специфічні ферменти.
Ген цих специфічних ферментів біодеградації часто розташовується
не в бактеріальній хромосомі, а в мобільному генетичному елементі,
такому як плазміда.
ДНК плазміди з
геном біодеградації
мРНК, яка кодує
ген деградації
Ферменти для
біодеградації
ксенобіотиків
Хромосома Плазміда
Клітина
8.
РНК
РНК (рибонуклеїнова кислота) -
лінійнійполімер нуклеотидів, до складу яких
входять залишок фосфорної кислоти, рибоза (на
відміну від ДНК, що містить дезоксирибозу) і
азотисті основи —аденін, цитозин, гуанін і урацил
(на відміну від ДНК, що містить тимін замість
урацилу).
На відміну від ДНК, яка існує у формі подвійної
спіралі, що складається з двох окремих молекул,
молекули РНК преважно одноланцюжкові.
Молекули РНК, в середньому, набагато коротше,
ніж молекули ДНК. Молекула РНК містить
декілька тисяч нуклеотидів, а молекула ДНК може
складатися з мільйонів нуклеотидів.
ДНКРНК
9.
Існує три класаРНК, які важливі для перетворення генної послідовності ДНК
на послідовність амінокислот в молекулі білка: матрична РНК, транспортна
РНК та рибосомальна РНК.
Матрична РНК, мРНК (синонім інформаційна РНК) —відповідає за перенесення
інформації про первинну структуру білків від ДНК до місць синтезу білків.
мРНК синтезується на основі ДНК в ході транскрипції.
Транскрипція
Спочатку ДНК клітини деспіралізується, і на одній із її ниток, як на матриці,
фермент ДНК-залежна РНК-полімераза за принципом комплементарності
формує полірибонуклеотидний ланцюг, який називається інформаційною або
матричною РНК (іРНК, мРНК). Принцип комплементарності: гуанін завжди
комплементарен цитозину, а аденін завжди комплементарен урацилу. Ця стадія
синтезу білка називається транскрипція (transcriptio - переписую).
ДНК (послідовність гену)
Послідовність мРНК
10.
Транспортні РНК (тРНК).У цитоплазмі клітини завжди існують транспортні
РНК. На одному з кінців вони мають триплет нуклеотидів (антикодон), а на
іншому - місце для з’єднання з відповідною амінокислотою (кодон). Кодон – це
послідовність з трьох нуклеотидів в мРНК, відповідна послідовності ДНК, що
кодує визначену амінокислоту. Для кожної амінокислоти існує своя тРНК.
Амінокислота ковалентно приєднується
до 3'- кінця молекули за допомогою
ферменту аміноацил-тРНК-синтетази.
Рибосомальні РНК (рРНК) — головний компонент рибосом.
11.
Однією зі стадійпроцесу біосинтезу білків є трансляція.
Трансляція
(translatio - перенесення). мРНК переноситься клітиною в цитоплазму, де
розташовані рибосоми. На рибосомах відбувається зчитування генетичного
коду мРНК і генетична інформація послідовності матричної (інформаційної)
РНК транслюєтся в послідовність амінокислот в білку.
Кожній амінокислоті відповідає своя тРНК, що має відповідний антикодон,
«відповідний» до кодону мРНК. Під час трансляції рибосома рухається
уподовж мРНК, у міру цього нарощується поліпептидний ланцюжок.
Процес трансляції можна поділити на чотири фази: активацію, ініціацію,
елонгацію та термінацію.
Синтез білка завжди починається з AUG-кодону, що також кодує метіонін.
Цей кодон зазвичай називають стартовим або ініциаторним.
Елонгація відбувається, коли чергова аміноацил-тРНК використовується для
збільшення поліпептидного ланцюжка. Термінація відбувається, коли
рибосома зустрічає стоп-кодон (UAA, UAG або UGA), для якого не існує
відповідної тРНК, при цьому відбувається звільнення поліпептидного
ланцюжка.
12.
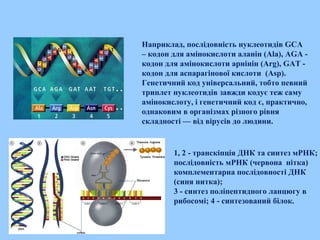
Наприклад, послідовність нуклеотидівGCA
– кодон для амінокислоти аланін (Ala), AGA -
кодон для амінокислоти арнінін (Arg), GAT -
кодон для аспарагінової кислоти (Asp).
Генетичний код універсальний, тобто певний
триплет нуклеотидів завжди кодує теж саму
амінокислоту, і генетичний код є, практично,
однаковим в організмах різного рівня
складності — від вірусів до людини.
1, 2 - транскіпція ДНК та синтез мРНК;
послідовність мРНК (червона нітка)
комплементарна послідовності ДНК
(синя нитка);
3 - синтез поліпептидного ланцюгу в
рибосомі; 4 - синтезований білок.
13.
Мінливість мікроорганізмів
Розрізняють двавиди мінливості мікроорганізмів:
•неспадкову або модифікаційну та
•спадкову або генотипну.
Модифікаційна мінливість полягає у зміні різноманітних властивостей
мікроорганізмів під впливом факторів навколишнього середовища, однак
вона не зачіпає генетичний апарат клітини спадково не передається.
При генотипній мінливості мікроорганізмів різноманітні ознаки бактерій
успадковуються та передаються нащадкам. Вона може розвиватись
внаслідок мутацій та рекомбінацій.
14.
Мутації
Мутація (лат. mutatio- зміна) – стійкі (тобто
такі, які можуть бути успадкованими) зміни в
послідовності нуклеотидів ДНК. Процес
виникнення мутацій дістав назву мутагенезу, а
організм зі змінами – мутанта.
За своїм походженням мутації поділяють на спонтанні та індуковані:
•спонтанні мутації відбуваються з частотою 105
-1012
без втручання
експериментатора, за, нібито, оптимальних умов існування мікробів. Вони
виникають внаслідок дії якихось не встановлених навколишніх факторів;
•індуковані мутації виникають під впливом дії на клітину мутагенних факторів.
Частота їх на декілька порядків вища, ніж спонтанних.
15.
За локалізацією мутаціїподіяють на:
• нуклеоїдні, які виникають в нуклеоїді клітини, та
•цитоплазматичні, що виникають у позахромосомних елементах спадковості –
плазмідах.
Мутації може бути:
• прямими, якщо їх прояв призводить до відхилення від ознак з так званого
дикого типу, і
•зворотними, якщо вони приводять до відновлення дикого типу.
Хімічні мутагени
Аналоги азотистих основ можуть включаються в молекулу ДНК і викликати
вставку некоректної нуклеотидної основи при реплікації.
Наприклад, бромураціл аналогічний за структурою тиміну, він включається в
ДНК як партнер аденіну, а потім переходить в енольну форму і впізнається
полімеразою як цитозин, що призводить до включення гуаніну замість аденіну.
Інші хімічні мутагени діють прямо на ДНК і викликають зміни в структурі
генів. Це деякі ароматичні речовини, важкі метали, азотиста кислота.
16.
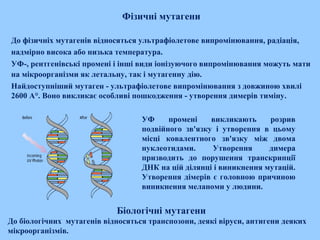
Фізичні мутагени
До фізичніхмутагенів відносяться ультрафіолетове випромінювання, радіація,
надмірно висока або низька температура.
УФ-, рентгенівські промені і інші види іонізуючого випромінювання можуть мати
на мікроорганізми як летальну, так і мутагенну дію.
Найдоступніший мутаген - ультрафіолетове випромінювання з довжиною хвилі
2600 А°. Воно викликає особливі пошкодження - утворення димерів тиміну.
Біологічні мутагени
До біологічних мутагенів відносяться транспозони, деякі віруси, антигени деяких
мікроорганізмів.
УФ промені викликають розрив
подвійного зв'язку і утворення в цьому
місці ковалентного зв'язку між двома
нуклеотидами. Утворення димера
призводить до порушення транскрипції
ДНК на цій ділянці і виникнення мутацій.
Утворення дімерів є головною причиною
виникнення меланоми у людини.
17.
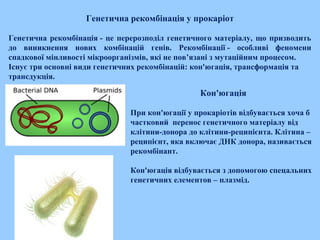
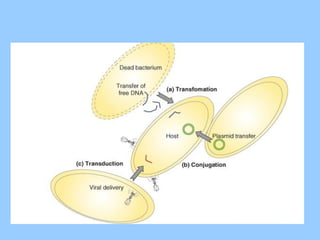
Генетична рекомбінація упрокаріот
Генетична рекомбінація - це перерозподіл генетичного матеріалу, що призводить
до виникнення нових комбінацій генів. Рекомбінації - особливі феномени
спадкової мінливості мікроорганізмів, які не пов’язані з мутаційним процесом.
Існує три основні види генетичних рекомбінацій: кон'югація, трансформація та
трансдукція.
Кон'югація
При кон'югації у прокаріотів відбувається хоча б
частковий перенос генетичного матеріалу від
клітини-донора до клітини-реципієнта. Клітина –
реципієнт, яка включає ДНК донора, називається
рекомбінант.
Кон'югація відбувається з допомогою спецальних
генетичних елементов – плазмід.
18.
У кишкової паличкиклітина-донор («чоловіча») має довгасту форму, клітина-
реципієнт («жіноча») - ізодіаметрічну. Клітка-донор утворює статеві ворсинки
(пілі), які притягують її до клітини-реципієнта і утворюють цитоплазматичні
канали. По цих каналах ДНК з клітини-донора переходить в клітину-реципієнт.
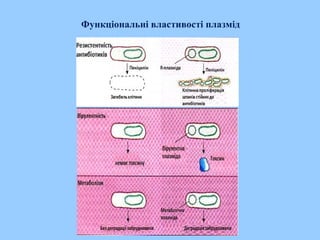
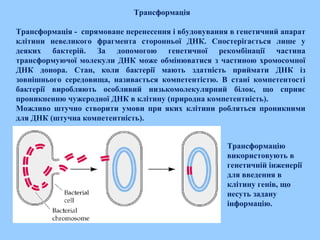
Трансформація
Трансформація -спрямоване перенесення і вбудовування в генетичний апарат
клітини невеликого фрагмента сторонньої ДНК. Спостерігається лише у
деяких бактерій. За допомогою генетичної рекомбінації частина
трансформуючої молекули ДНК може обмінюватися з частиною хромосомної
ДНК донора. Стан, коли бактерії мають здатність приймати ДНК із
зовнішнього середовища, називається компетентістю. В стані компетентості
бактерії виробляють особливий низькомолекулярний білок, що сприяє
проникненню чужеродної ДНК в клітину (природна компетентність).
Можливо штучно створити умови при яких клітини робляться проникними
для ДНК (штучна компетентність).
Трансформацію
використовують в
генетичній інженерії
для введення в
клітину генів, що
несуть задану
інформацію.
21.
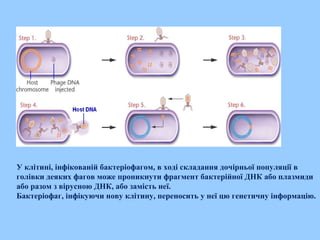
Трансдукція
Трансдукція —передача генетичногоматеріалу від однієї клітини (донора) до
іншої (реципієнтові) відбувається за допомогою віруса (бактеріофага у
випадку бактерій), що призводить до зміни спадкових властивостей.
22.
У клітині, інфікованійбактеріофагом, в ході складання дочірньої популяції в
голівки деяких фагов може проникнути фрагмент бактерійної ДНК або плазмиди
або разом з вірусною ДНК, або замість неї.
Бактеріофаг, інфікуючи нову клітину, переносить у неї цю генетичну інформацію.
24.
Генетична інженерія
-це знаннята методи створення організмів in vitro з новими комбінаціями
спадкових ознак, одержання конкретних генів, що визначають ту чи іншу
ознаку клітини або організму.
Методи генної інженерії дають змогу поєднувати генетичну інформацію
найрізноманітніших організмів, наприклад, з'єднуючи гени тварин з
генами мікробів або рослин.
Генна інженерія дозволяє створювати мікробні культури з підвищинним
синтезом цінних метаболітів або з підвищеною мікробною активністю.
25.
Одержання рекомбінантних молекул.Ферменти рестрикції.
Для одержання рекомбінантних молекул
ДНК застосовуються ферменти
рестрикції (рестриктази), які здатні роз
пізнавати специфічні послідовності основ
у дволанцюговій молекулі ДНК, а потім
розщеплювати такі молекули на
фрагменти. Поділ подвійного ланцюга ДНК.
Одноланцюгові комплементарні кінці двох ДНК називають "липкими кінцями",
тому що вони з'єднуються внаслідок комплементарного спарювання азотистих
основ. Вони полегшують вставку чужорідної ДНК у векторну.
Вектори для переносу генів і фрагментів ДНК
Векторами в генній інженерії служать плазміди, фагі, віруси, мобільні елементи.
Існує два типи плазмід — однокопійні (на клітину припадає 1 молекула плазмідної
ДНК) і мультикопійні (на одну клітину припадає 10—20 плазмідних геномів).
Окремі плазміди можуть накопичуватися у величезних кількостях — до 1 тис.
плазмід на одну клітину. Такі плазміди використовуються для клонування
векторів.
26.
Можна поєднувати відрізкиДНК,
отримані з різних джерел, і
створювати комбінації генів в одній
довгій молекулі. Для з'єднання
ділянок застосовують фермент лігазу.
Комбінуючи різні рестриктази і
лігази, можна розрізати нитку ДНК у
різних місцях і одержувати
рекомбінантні молекули (наприклад,
плазмідну ДНК з вмонтованим чужим
геном).
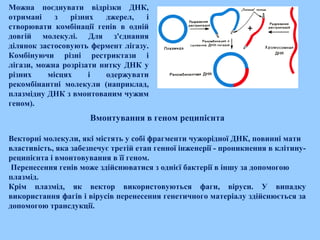
Вмонтування в геном реципієнта
Векторні молекули, які містять у собі фрагменти чужорідної ДНК, повинні мати
властивість, яка забезпечує третій етап генної інженерії проникнення в клітину
реципієнта і вмонтовування в її геном.
Перенесення генів може здійснюватися з однієї бактерії в іншу за допомогою
плазмід.
Крім плазмід, як вектор використовуються фаги, віруси. У випадку
використання фагів і вірусів перенесення генетичного матеріалу здійснюється за
допомогою трансдукції.
27.
Гени біодеградаціх ксенобіотиківбули ізольовано і вмонтовані у плазміди.
Штами, що містять такі плазміди можуть бути використані для очищення
грунту забрудненому ксенобіотиками.
Головна проблема забезпечення стабільності плазмід у таких штамів.
Практичне значення генетики бактерій
Генетична інженерія знайшла широке практичне застосування в різних
галузях науки, техніки, медицини, фармацевтичної промисловості,
біотехнології, сільського господарства.
Завдяки застосуванню генетичних методів, одержано високоактивні штами
бактерій, грибів, актиноміцетів, дріжджів, які продукують у 2001000 разів
більше амінокислот, органічних кислот, ферментів, вітамінів, кормового білка
порівняно з вихідними, а також вакцинні штами мікроорганізмів та вірусів.
Для посилення синтезу певних продуктів можливо використовувати
клонування генів процес, що включає виділення й ампліфікацію (дублювання
великої кількості) окремих генів у реципієнтних клітинах.
Рекомбінантні штами для деградації ксенобіотиків
28.
Використання різноманітних мутагенів(ультрафіолетове та радіоактивне
опромінення, хімічні речовини) дозволило створити мутантний штам гриба
Penicillium chryzogenum, який у дикому стані продукував 100 од/мл
пеніциліну, а після направленої селекції 10000 од/мл.
На даний час методи генетичної інженерії з успіхом використовують для
одержання бактеріальних штамів продуцентів біологічно активних
сполук, у тому числі гормонів (інсуліну, гормону росту – соматотропіну
недолік цього гормону призводить до карликовості), противірусного
препарату інтерферону та ін.
Кишкова паличка –продуцент інсуліну.
Про високу ефективність генноінженерних
методів одержання хімічно чистих біологічно
активних речовин свідчить такий факт. Щоб
одержати 5 мг соматотропіну, було
використано мозок 500 000 овець протягом 5
років, в той час як аналогічну кількість
гормону дають 9 л суспензії кишкової
палички.
Біоінженерні бактерії використовуються для оздоровлення рослин. Так, у
бактерій, що утворюють симбіоз з коренями кукурудзи, були введені, гени (від
інших бактерій), що кодують токсин для шкідливих комах.