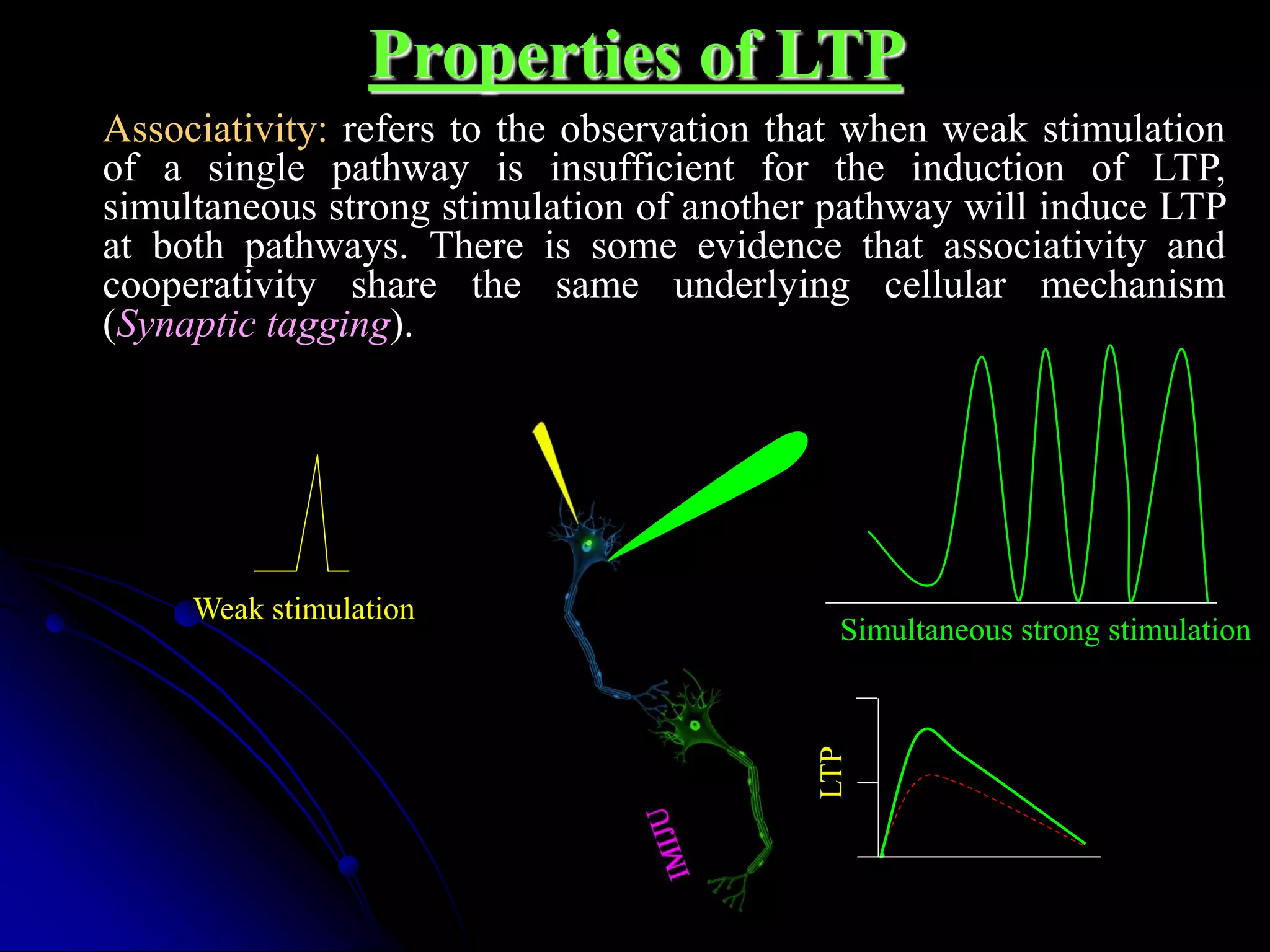
Weak stimulation
No LTP
Strong stimulation of other pathway
LTP at both pathways
Associativity
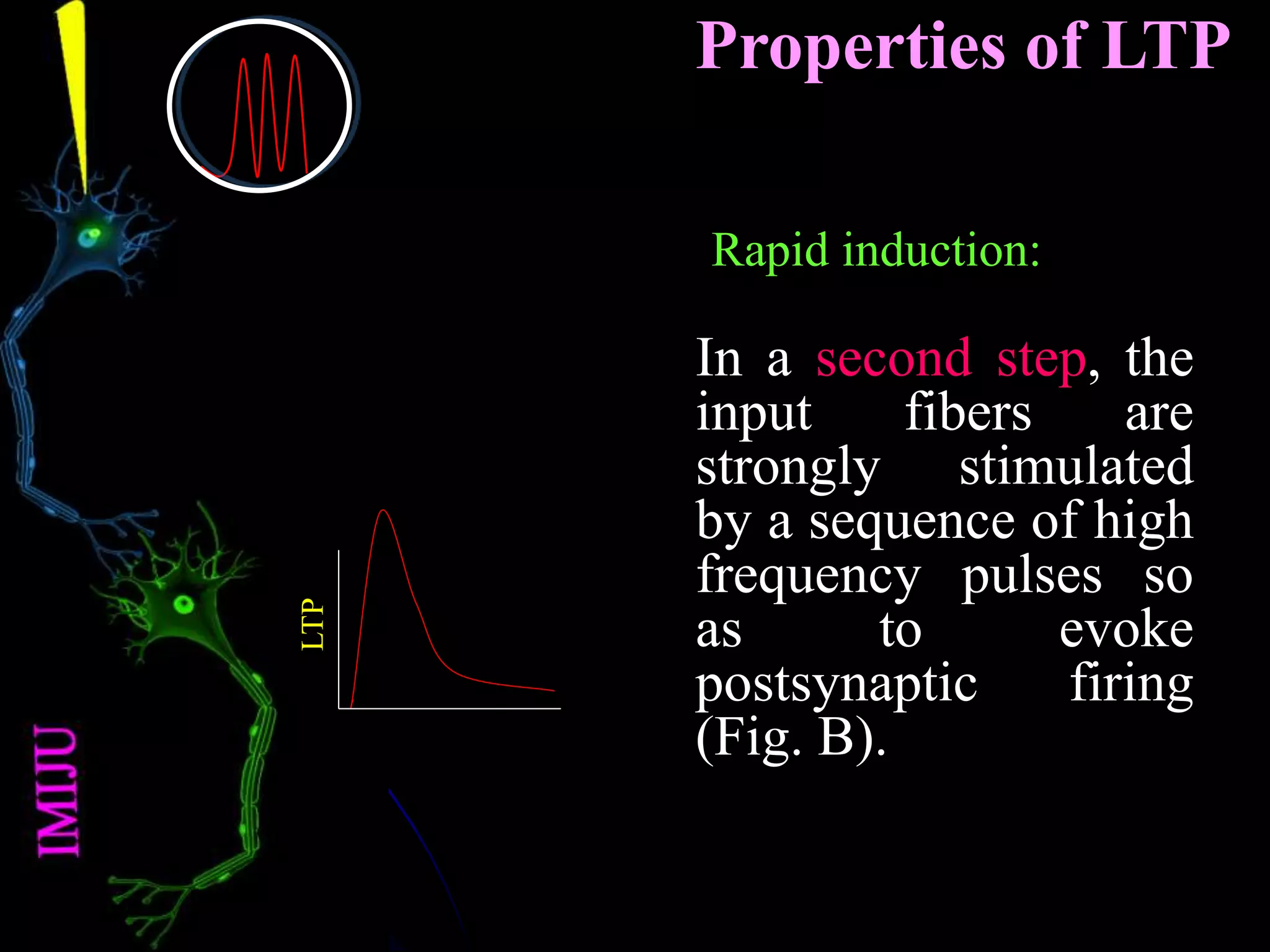
�Properties of LTP
Input specificity: Once induced, LTP at one synapse is not
propagated to adjacent synapses; rather LTP is input specific.
LTP induced synapse
No LTP at other synapses
Input specificity