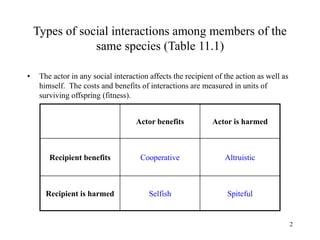
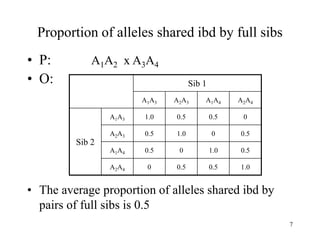
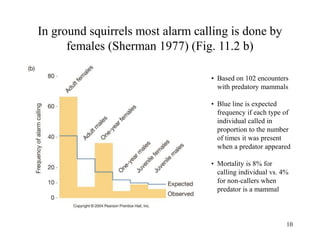
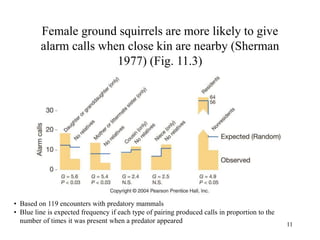
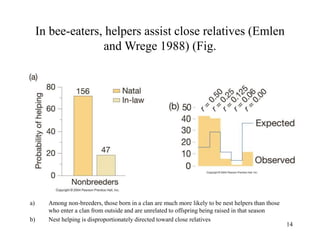
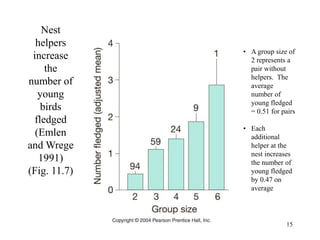
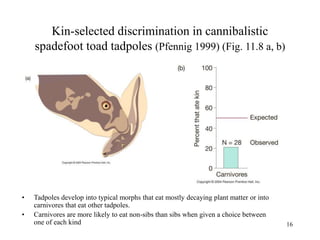
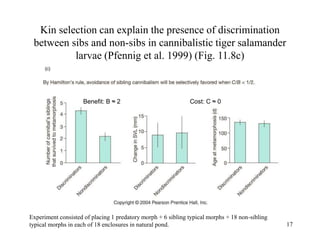
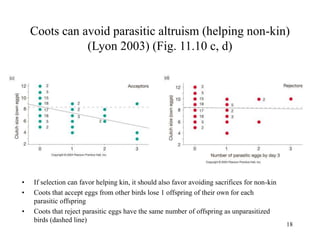
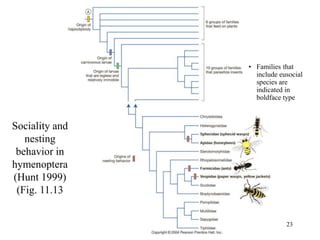
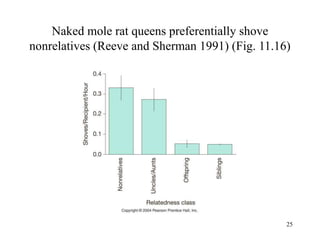
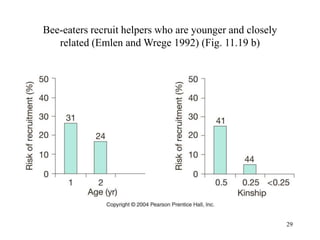
Kin selection and social behavior can be explained through Hamilton's rule of kin selection (Br-C>0). Altruistic behaviors that decrease an individual's direct fitness can increase if they provide sufficient benefits to the reproductive success of genetic relatives. This explains phenomena like alarm calling in ground squirrels, where females are more likely to call when close kin are present. Haplodiploidy in hymenoptera may have predisposed the evolution of eusociality as workers are more closely related to sisters (r=3/4) than to their own offspring (r=1/2). Parent-offspring conflict can arise when parents and offspring differ in how they value the costs and benefits of continued parental care















![27
Parent – offspring conflict occurs because parents and offspring value the
costs of parental care differently (Trivers 1985) (Fig 11.18)
• As offspring grow, the benefit/cost ratio for the parent declines. Benefit (B) is
measured in terms of increased survival of offspring receiving parental care; cost (C) is
measured in terms of lost future offspring due to continued parental care. From
parent’s point of view, it should stop giving parental care when B/C declines to 1.
• But, the offspring devalues the cost to the parent by 1/2 because lost future full-sibs are
related to the offspring by r = 1/2. Therefore, the offspring wants parental care to
continue until the B/C ratio for the parent is 1/2 - fig. (a) [(or 1/4 if future offspring are
half-sibs - fig. (b)]](https://image.slidesharecdn.com/kinselection-240212042305-c15ca6ec/85/Kin-selection-with-respect-to-animals-ppt-27-320.jpg)













![Neural_control_of_animal_behaviour[1]789.pptx](https://cdn.slidesharecdn.com/ss_thumbnails/neuralcontrolofanimalbehaviour1789-230207172442-5de82c6f-thumbnail.jpg?width=640&height=640&fit=bounds)

























































