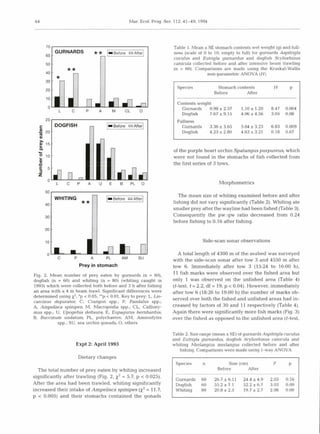
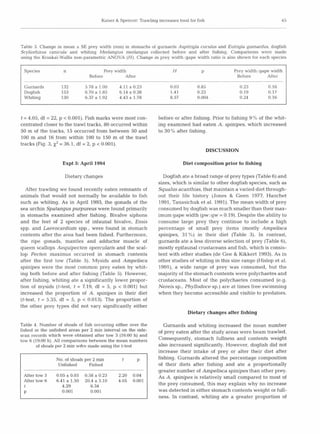
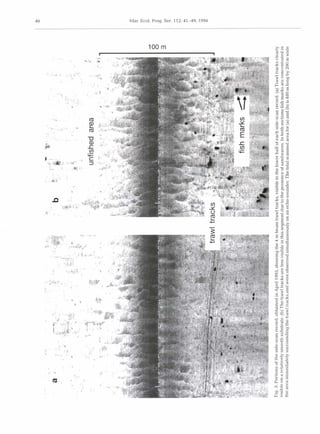
This study examined the diets of four fish species to determine if they feed on animals damaged by beam trawling. The researchers conducted beam trawling experiments off the coast of Wales and analyzed the stomach contents of fish caught before and after trawling. They found that two species, gurnards and whiting, significantly increased their food intake after an area was trawled, eating more of certain prey like amphipods that were likely disturbed by the trawling. Observations with sonar also detected more fish concentrated near the trawl tracks compared to untrawled areas. The results suggest these fish rapidly migrate into recently trawled areas to scavenge on damaged or disturbed animals, potentially altering community structure with































![Journal Micropalaeontology:Crittenden 1982-jm1-1-23[1]. Rotalline foraminifer...](https://cdn.slidesharecdn.com/ss_thumbnails/crittenden-1982-jm1-1-231-170321113030-thumbnail.jpg?width=640&height=640&fit=bounds)






























































