Download to read offline




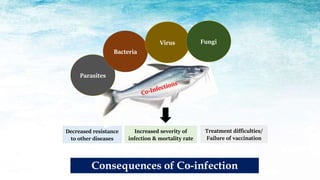
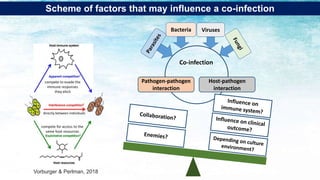

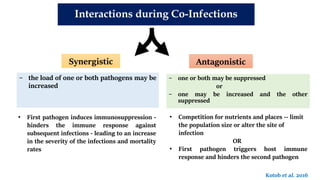
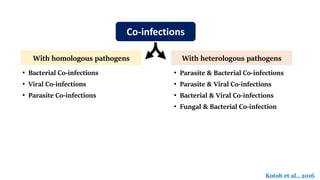


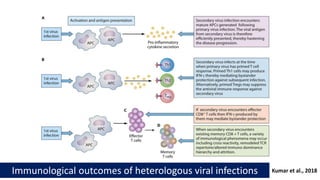

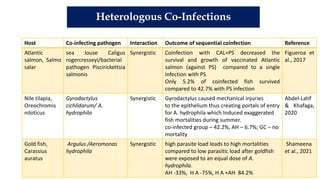
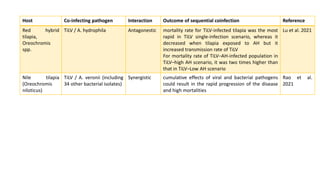
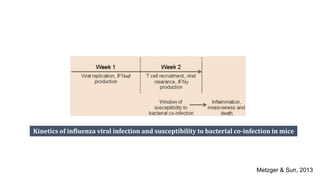
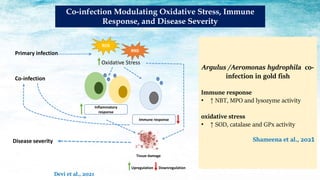

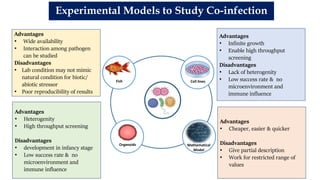



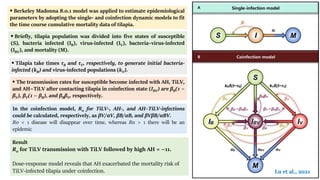
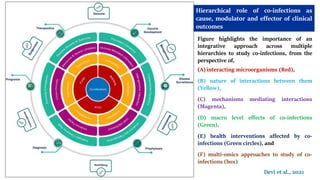





The document discusses the significant economic losses in aquaculture due to diseases and co-infections in farmed fish, highlighting that co-infections often lead to increased mortality and treatment challenges. It explains the dynamics of host-pathogen interactions during concurrent infections and presents various case studies illustrating the effects of specific co-infections on different fish species. The conclusion emphasizes the need for integrated approaches to study co-infections and their implications on fish health and aquaculture productivity.









































































