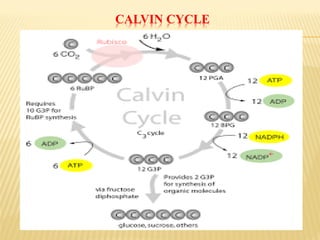
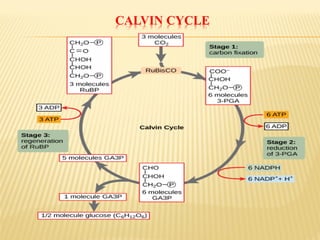
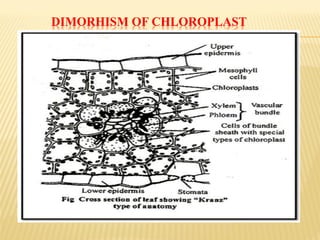
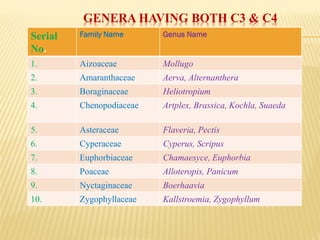
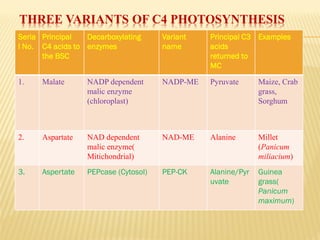
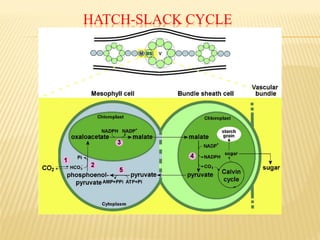
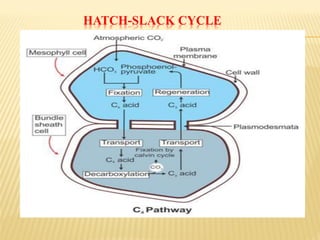
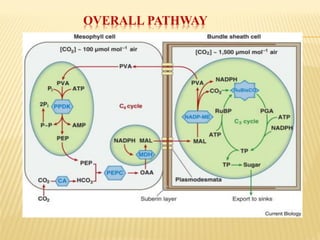
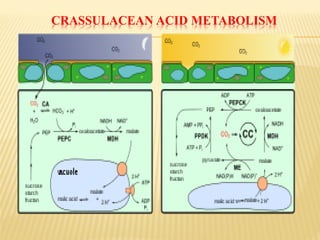
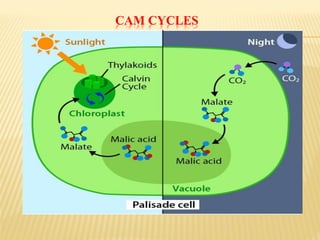
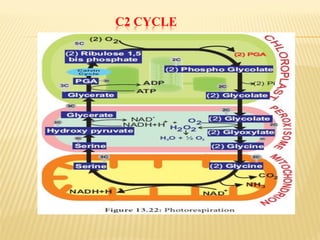
The document discusses carbon assimilation in plants, detailing the major pathways including the Calvin cycle (C3), C4 pathway, and Crassulacean Acid Metabolism (CAM). It explains the biochemical processes involved in photosynthesis, highlighting the differences between C3, C4, and CAM plants in terms of their carbon fixation mechanisms and environmental adaptations. Furthermore, it describes the unique structures and functions of chloroplasts within these plant types, emphasizing the significance of these pathways in maximizing efficiency under varying environmental conditions.





























































![Biosynthesis of auxin[1]](https://cdn.slidesharecdn.com/ss_thumbnails/biosynthesisofauxin1-170505044723-thumbnail.jpg?width=640&height=640&fit=bounds)
































































