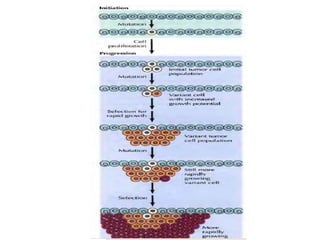
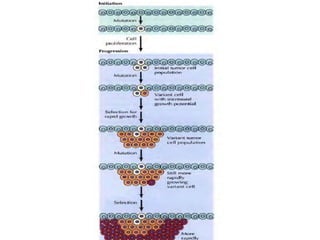
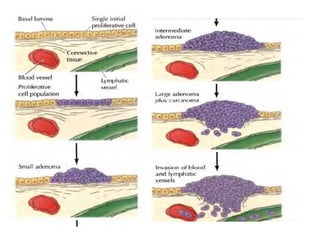
Cancer develops through a multistep process where cells accumulate mutations that allow uncontrolled growth. Normal cells undergo programmed cell death and have contact inhibition, limiting factors that cancer cells evade. Cancer cells also secrete factors like proteases and angiogenic growth factors that promote invasion and metastasis. Overall, cancer arises through genetic and environmental factors that disrupt the normal cell cycle, proliferation, differentiation and survival mechanisms.