
Recommended
More Related Content
What's hot
What's hot (20)
Viewers also liked
Viewers also liked (20)
Similar to photomorphogenesis, photosensors, light signal transduction
Similar to photomorphogenesis, photosensors, light signal transduction (20)
More from Kevin Folta
More from Kevin Folta (8)
Recently uploaded
Recently uploaded (20)
photomorphogenesis, photosensors, light signal transduction
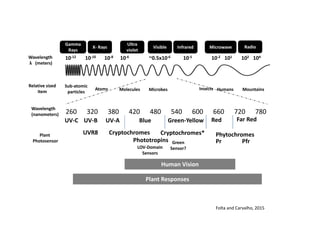
- 1. Wavelength λ (meters) 10‐12 10‐10 10‐8 ~0.5x10‐6 10‐5 10‐2 102 Gamma Rays X‐ Rays Ultra violet Visible Infrared Microwave Radio Sub‐atomic particles Atoms Molecules Microbes Insects 10‐6 104 101 Humans Mountains Relative sized item Wavelength (nanometers) 260 320 380 420 480 540 600 660 720 780 UV‐C UV‐B UV‐A Blue Green‐Yellow Red Far Red UVR8 Cryptochromes Cryptochromes* Phytochromes Pr PfrPhototropins LOV‐Domain Sensors Green Sensor? Plant Photosensor Human Vision Plant Responses Folta and Carvalho, 2015
- 2. Borthwick’s Experiment in Grand Rapids Lettuce (1954)
- 3. -synthesized as Pr in darkness -converted by red light (max= 666 nm) to Pfr -Pfr is biologically active form -Far-red light (730 nm) converts to Pr
- 4. Early Developmental Effects of Phytochromes Phytochromes regulate (100’s of processes described) -- stem elongation -- cotyledon expansion -- chloroplast development (greening) -- apical hook opening -- gene expression Microarray analyses show that the early effects of phytochromes are to induce the expression of transcription factors that then alter the expression of genes involved in photomorphogenic development (Tepperman et al., 2001).
- 5. Shade avoidance responses --- Induced under FR enriched circumstances ---EOD-FR(End of Day Far Red) can induce a similar response ---Can be induced by shade or light reflected from neighboring plants (“density sensor?”) ---Controlled by Type II phytochromes ---Plants possess long petioles, longer stems and unusually elongated organs in general. Regulation of Flowering time ---phyB mutant shows an early flowering in LD (long day 16H/8D) condition. ---phyA mutant shows an late flowering in Extended SD (8L/8 FR-enrich/8D) condition. Long-term Roles for Phytochrome in Regulation of Plant Physiology
- 6. ENTER ARABIDOPSIS Koornneef M., Rolff E. and Spruit C.J.P. (1980) Genetic control of light- inhibited hypocotyl elongation in Arabidopsis thaliana (L.) Heynh. Z. Pflanzenphysiol. 100: 147-160. hy Mutants normal phy? hy1 yes hy2 yes hy3 no hy4 yes hy5 yes additional hy’s isolated later hy8 no Mutation in heme oxygenase (Davis et al., 2001) mutation in heme oxygenase mutation of phytochrome B mutation of cryptochrome1, a photolyase-like protein mutation in a B-zip transcription factor mutation of phytochrome A (Dehesh et al, 1993) Would turn out to encode:
- 7. Has topology similar to histidine kinases, yet exhibits ser-thr kinase activity. phytochrome binds ATP diverged from prokaryotic orthologs after duplication of kinase transmitter domain (Yeh and Lagarius, 1998). BUT IS IT A KINASE?
- 8. Type I phytochrome : photo-labile (phyA) Type II phytochrome : photo-stable (phyB, phyC, phyD, phyE) Two Major Types of Phytochrome: B, R, FR PrA PfrA Dark reversion Destruction PrB PfrB Dark reversion R FR FR
- 9. The phy chromophore is a tetrapyrrole that is reliant on heme oxygenase for its synthesis; mutants in heme oxygenase 1 exhibit phy-like phenotypes. The phenotypes can be rescued by exogenous application of biliverdin (Parks and Quail, 1991). increasing BV Brian Parks, circa 2000 and Brian’s homespun Dobsonian mount telescope, circa 2003.
- 10. Parks and Quail, 1993 Peter Quail hy8 and hy1 mutants exhibit specific inability to sense far-red; hy8 mutants sense red light just fine…
- 11. Parks and Quail, 1993 The phenotypes of the phytochrome mutants defined as hy loci. Growth is unaffected in darkness. hy3 and hy1 are impaired in red light sensing hy8 and hy1 are impaired in far-red light sensing
- 12. Red – Import within minutes Import of phyA to nucleus by red light also occurs within minutes; followed by disappearance of the protein. A phyB::GFP fusion protein is strongly detectable in the nucleus of transgenic tobacco cells beginning after three hours (Gil et al., 2000).
- 13. Far-red– import after 1.5 h; retained throughout time course (Hisada et al, 2000). phyA effects FR physiology with a similar time course; protein constantly detectable in WT background. parks and spalding, 1999
- 14. Null phyB mutants are tested for complementation using the GR fusions. The results indicate that both light and nuclear import are required for full restoration of phyB activity.
- 15. Interaction between PIF3 and PHYB is dependent upon state of PHYB PIF3 and PHYB only interact if phyB is in Pfr state. Ni et al., 1999
- 16. PIF3 Interaction with Promoter is Sequence Specific, and is Mediated Through the DNA Binding Domains Martinez-Garcia et al., 2000
- 17. Martinez-Garcia et al., 2000 Binding of phyB to the Promoter Requires PIF3, the phyB Chromoprotein in the Pfr Form, and PIF3 Domains Outside of the DNA-Binding Domains
- 18. deLucas et al., Nature 2008
- 22. Claus Schwechheimera, and Björn Christopher Willigea Curr op plant bio, 2008
- 23. PKS, NDPK2
- 24. Cryptochromes The sequence of Hy4 was reported in 1993, and it was similar to DNA photolyase (Ahmad and Cashmore 1993, Sancar 1994) yet has no photolyase activity. Re-designated “Cryptochrome 1” (Lin et al., 1995) Involvement in circadian rhythms; led to discovery of animal crys (Cashmore, 2003) Many plant responses were not R-FR reversible and had action spectra with peaks in the blue and near-UV. There must be a BL receptor(s). Due to their elusive nature, Gressel (1977) described BL receptors as “cryptochromes”. hy4 mutants showed a lack of hypocotyl growth inhibition under blue light,but were normal under red and far-red. Dark BL Wild-type BL cry1 adapted from Neff and Chory, 1998
- 25. Photoactivation of cryptochromes is (likely) similar to what is observed in photolyases. Both a folate (MTHF) and a flavin serve as chromophores in photolyase, and these two compounds purify with cry when it is expressed in E.coli (Malorta et al., 1995). model of activity in photolyase Cashmore, 2003.
- 26. cry2 mutants The Cry2 gene was identified in the genome by homology to Cry1 (Jarillo et al., 1996). The cry2 mutants were isolated as seedlings not expressing CRY2 protein. The mutants flowered late and were shown to be allelic to other flowering-time genes (below; Guo et al., 1998). cry2 mutants lack a conspicuous end-point phenotype under high- fluence blue light (right; Mockler et al., 1998) 50 µmol m-2 s-1 5.5 µmol m-2 s-1 0.6 µmol m-2 s-1
- 27. CRY2 is phosphorylated in a light-dependent manner (Shalitin et al., 2002) 1. Test for light-dependent phosphorylation -- incubate with γ32P-ATP 2. Test to verify presence of CRY2 in same conditions 3. Specific gel buffer/ionic conditions allow resolution of the phosphorylated form of the protein. Again, the phosphorylated form (P) is detectable only after BL exposure. P
- 28. The timing of CRY1 and CRY2 phosphorylation generally coincides with CRY2’s influence on early growth suppression. Shalitin et al., 2002, 2003 Folta and Spalding, 2001 CRY2 phosphorylation CRY1 phosphorylation
- 29. CRY2 accumulates in cop mutants The phosphorylated form accumulates in cop1– and both the phosphorylated and unphosphorylated forms disappear in WT. Propose a mechanism.
- 30. Xing Wang Deng, Yale COP Mutants First isolated by Deng et al., 1991 --Constitutive Photomorphogenic phenotype: expanded cotyledons, short hypo- cotyls, light-regulated gene expression patterns in darkness 1996 Mayer et al show that COP1 mutation affects expression of many genes OLD SLIDE REVISITED!
- 31. The hy5 mutant – long hypocotyl under light conditions, particularly blue (Koorrneef et al., 1980). Encodes a B-zip transcription factor that is presumably a positive regulator of photomorphogenesis. HY5 accumulates rapidly in light and is not as detectable upon transfer of plants to darkness fluence rate Plants grown for days in light show different levels of HY5– HY5 level correlates with advanced photomorphogenic development. Osterlund et al., 2000 How does HY5 regulate photomorphogenesis? Is it simply present only in light and acting as a positive regulator? Is it more complex? It seems to be acting in a manner that is antagonistic to COP1….
- 32. HY5 mRNA accumulates in both dark and light conditions, but the protein is detected only in light. Osterlund et al., 2000 Is this another example of ubiquitin-mediated proteolysis? How can this be tested?
- 33. 1. Test using genetic tools – cop1 mutants 2. Test using pharmacological tools – proteosome inhibitors Proteosome inhibitors allow increased accumulation of HY5. mRNA levels are the same in cop and WT plants in response to light treatment, yet protein levels are very different. This strongly suggests that COP1 has a role in regulating the accumulation of HY5, a positive regulator of photomorphogenesis. Osterlund et al,. 2000
- 34. These results support a model where HY5, a positive regulator of photomorphogenesis is degraded in darkness via COP1 and the proteosome. Osterlund et al., 2000
- 35. The C-Terminus of cry1 Regulates Photomorphogenesis Ectopic overexpression of the CRY1 C-terminal extension results in a constitutive-photomorphogenic phenotype. Cashmore, 2003 Is cry functioning through a mechanism involving COP1?
- 36. Does COP1 Interact with CRY? Yeast 2-hybrid assay between COP1 truncations and CCT2 (CRY2 c-terminus) Conclusions: The WD repeat domain is necessary and sufficient for interaction, yet binding is strongly enhanced by the coiled-coil and Zn binding domains. CCT2-GUS interacts similarly. Coimmunoprecipitation – Using anti-CRY2, COP1 can be coimmunoprecipitated, demonstrating likely interaction in vivo. Wang et al., 2001
- 37. Hellmann and Estelle, 2002 Proposed Model for Cryptochrome Function – COP1, CRY, HY5 Interaction to Regulate Degradation of HY5
- 38. Phototropism Noted by Darwin in observations of dark-grown canary grass Colodny-Went model proposed that auxin would be secreted from the tip and light would guide a lateral gradient of auxin to accumulate, leading to differential growth. A split in the plant biology community– phototropism just a phy response vs a specialized BL receptor
- 39. Propose a Forward Genetic Screen - what might you find? photoreceptor(s)– the actual molecule that receives the photon signaling mutants– non-redundant genes required for transduction of the signal from activated photoreceptor to target genes. response mutants – those that have impaired cell wall expansion, auxin transporters required for differential auxin redistribution… etc. ? upstream elements – possibly molecules required for photoreceptor synthesis and/or function.
- 40. phot1- BL-activated, membrane-associated, autophosphorylating ser/thr kinase with two LOV domains (Christie and Briggs, 1998). NPH3 - novel protein with possible BTB-POZ transcription activation domains. NPH3 functions downstream of phot1, possibly participating as a scaffold protein in the phosphorelay. RPT2 – deficient in root phototropism, very similar to NPH3 . NPH4- the same gene as ARF7 (Harper et al., 1999). The phototropic mutant loci would be shown to encode:
- 41. Phototropins Regulate: Phototropism (Huala et al., 1997; Christie et al., 1998) Chloroplast accumulation and avoidance (Kagawa, 2001; Jarillo, 2001) Primary hypocotyl growth inhibition (0-30 min; Folta and Spalding, 2001, Folta et al., 2003) Stomatal opening (Kinoshita et al., 2001) Leaf expansion (Sakamoto and Briggs, 2002) Blue-light destabilization of specific mRNA’s (Folta and Kaufman, 2003) Solar tracking?
- 42. Phototropin – The Phototropism Photoreceptor Mediates responses to blue and near UV light. Binds two flavin mononucleotide molecules as chromophore (Christie et al., 1998). phosphorylation.. signaling or desensitization? Palmer et al., 1993
- 43. Phot Topology Contains two specialized PAS domains called LOV domains (light, oxygen, voltage) and a kinase domain (Christie and Briggs, 2001). The protein can autophosphorylate on many sites (Salomon et al., 2003). PHOT1 and PHOT2 share 60% sequence homology. ,
- 44. A Summary of Phot-Mediated Responses phototropism chloroplast relocation stomatal aperture low fluence-rate blue high fluence-rate blue Christie and Briggs, 2001 Sakai et al., 2001 accum avoid
- 45. Chloroplast accumulation and avoidance responses LFR BL HFR BL Treatment of leaf surfaces with low-fluence-rate blue light results in migration of chloroplasts to the anticlinal surfaces of the leaf, perpendicular to incident light. Irradiation with high-fluence-rate blue light results in migration of chloroplasts to the periclinal cell surfaces, parallel to incident light. This is important because it balances photosynthetic potential against photoinhibition (Kasahara et al., 2002).
- 46. phot2 (aka. cav1 and npl1) cav mutants obtained from forward genetic screen for plants that failed to show chloroplast avoidance. Gene mapped to a region containing NPL1, an NPH1 (phot1)- like gene. A reverse genetic screen for NPL1 resulted in plants that failed to relocate chloroplasts.
- 47. Photoreceptors of Adiantium Adiantium contains 2 phototropins, and shows normal phototropism and chloroplast accumulation responses to BLUE LIGHT… But the rap2 mutant does not exhibit chloroplast accumulation or curvature in response to red light. -- Red light mediates phototropism and chloroplast accumulation in Adiantium via a hybrid phy-phot photoreceptor.
- 48. Cryptochrome mysteries to be resolved: What is the mechanism of cry1 activation? How do redox changes affect signaling? cry1 and cry2 function within seconds of irradiation at the membrane, a process that is required for inhibition of stem growth (Folta and Spalding, 2001). Do cry’s have a signaling mechanism independent of COP1? cry interaction with phy, complete with cross phosphorylation has been reported (Ahmad et al., 1997; Mas et al., 2000; Jarillo et al., 2000).
- 49. Integrated Scheme of cry and phy function Quail, April 2002 What has changed already? Don’t memorize this for the exam! It may be very different in a year!