Mutia Rizqa Firdiah (25010112140145 kls B 2012) Biomol
•Download as DOCX, PDF•
0 likes•650 views

Genom Prokariot dan Eukariot

Recommended
Recommended
More Related Content
What's hot
What's hot (20)
Similar to Mutia Rizqa Firdiah (25010112140145 kls B 2012) Biomol
Similar to Mutia Rizqa Firdiah (25010112140145 kls B 2012) Biomol (20)
More from Mutia Rizqa Ofir
More from Mutia Rizqa Ofir (14)
Mutia Rizqa Firdiah (25010112140145 kls B 2012) Biomol
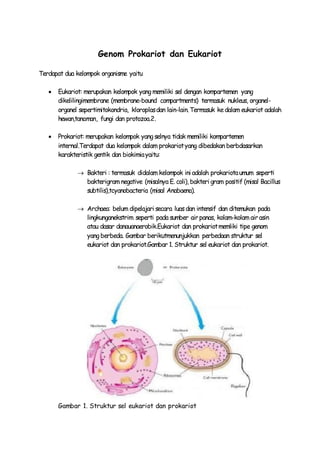
- 1. Genom Prokariot dan Eukariot Terdapat dua kelompok organisme yaitu Eukariot: merupakan kelompok yang memiliki sel dengan kompartemen yang dikelilingimembrane (membrane-bound compartments) termasuk nukleus, organel-organel sepertimitokondria, kloroplas dan lain-lain. Termasuk ke dalam eukariot adalah hewan,tanaman, fungi dan protozoa.2. Prokariot: merupakan kelompok yang selnya tidak memiliki kompartemen internal.Terdapat dua kelompok dalam prokariot yang dibedakan berbdasarkan karakteristik gentik dan biokimia yaitu: Bakteri : termasuk didalam kelompok ini adalah prokariota umum seperti bakterigram negative (misalnya E. coli), bakteri gram positif (misal Bacillus subtilis),tcyanobacteria (misal Anabaena). Archaea: belum dipelajari secara luas dan intensif dan ditemukan pada lingkunganekstrim seperti pada sumber air panas, kolam-kolam air asin atau dasar danauanaerobik.Eukariot dan prokariot memliki tipe genom yang berbeda. Gambar berikutmenunjukkan perbedaan struktur sel eukariot dan prokariot.Gambar 1. Struktur sel eukariot dan prokariot. Gambar 1. Struktur sel eukariot dan prokariot
- 2. A. Genom Eukariot Genom manusia merupakan model yang baik bagi genom eukariot secara umum. Genom nucleareukariotik memiliki molekul DNA linear yang terdapat di dalam kromosom. Semua eukariot juga memiliki genom yang lebih kecil yang berbentuk sirkular yaitu genom mitokondria. Pada tumbuhan, terdapat genom lain yaitu genom kloroplas.Walaupun struktur dasar eukariot mirip tetapi satu hal penting yang sangat berbeda adalah ukuran genom. Genom eukariot yang terkecil berukuran kurang dari 10Mb panjangnya. Sedangkan genom yang terbesar berukuran lebih dari 100 000 Mb. Variasi ukuran genom dapatdilihat pada tabel di bawah ini. Tabel 1. Ukuran genom eukariot (Brown, 2002) Spesies Ukuran genom (Mb) Fungi Saccharomyces cerevisiae 12.1 Aspergillus nidulans 25.4 Protozoa Tetrahymena pyriformis 190 Invertebrates Caenorhabditis elegans 97 Drosophila melanogaster 180 Bombyx mori (silkworm) 490 Strongylocentrotus purpuratus (sea urchin) 845 Locusta migratoria (locust) 5000 Vertebrates Takifugu rubripes (pufferfish) 400 Homo sapiens 3200 Mus musculus (mouse) 3300 Plants Arabidopsis thaliana (vetch) 125 Oryza sativa (rice) 430 Zea mays (maize) 2500 Pisum sativum (pea) 4800 Triticum aestivum (wheat) 16 000 Fritillaria assyriaca (fritillary) 120 000
- 3. Seperti terlihat pada tabel, ukuran genom bervariasi dan berhubungan dengan kekompleks anorganisme. Eukariot yang lebih sederhana seperti fungi memiliki genom yang paling kecil, daneukariot yang lebih tinggi seperti vertebrata dan tanaman berbunga memiliki genome yang lebihbesar. Hal ini mungkin terlihat masuk akal, karena kompleksitas organism diharapkan berhubungan dengan jumlah gen dalam genom – eukariot yang lebih tinggi memerlukan genomyang lebih besar untuk mengakomodasi gen ekstra. Tetapi korelasi ini jauh dari sempurna, jika korelasinya baik, maka genom nuklear yeast S. cerevisiae, yang berukuran 12 Mb adalah 0.004kali ukuran genom nuklear manusia, akan mengandung 0.004 × 35 000 gen yaitu hanya 140.Padahal kenyataannya genom S. cerevisiae mengandung 5800 gen.Tidak adanya korelasi antara kompleksitas suatu organism dengan ukuran genomnya, disebutsebagai C-value paradox. Jawabannya sederhana yaitu: tempat disiapkan di genom organisme yang kurang kompleks karena gen terpak bersama. Genom S. cerevisiae yang sekuensnya selesaidikerjakan tahun 1996 menggambarkan hal ini seperti terlihat pada gambar di bawah ini.
- 4. Gambar 2. Perbandingan genome manusia, yeast, lalat buah, jagung dan E. coli. (A) Segmen 50-kb dari lokus reseptor β T-cell manusia.(B) Saccharomycescerevisiae (chromosome III) (C)Drosophila melanogaster (D) jagung dan (E) E. coli K12Pada gambar di atas, segmen 50-kb dari genom manusia dibandingkan dengan segmen 50-kbgenom yeast. Segmen genom yeast yang berasal dari kromosom III memiliki karakteristik sebagai berikut: Memiliki lebih banyak gen dibandingkan segmen pada manusia. Daerah pada kromosom III yeast ini mengandung 26 gen yang mengkode protein dan dua yangmengkode transfer RNA (tRNA), molekul non-coding RNA terlibat dalam pembacaankode genetic selama proses sintesis protein. Relatif sedikit gen yeast yang discontinuous. Pada segmen kromosom III ini, tidak adagen yang discontinuous. Dalam keseluruhan genom yeast hanya terdapat 239 introns,dibandingkan dengan lebih dari 300 000 pada genom manusia. Terdapat lebih sedikit genome-wide repeats. Bagian kromosom III ini mengandungelemen sebuah repeat tunggal long terminal (LTR) element, disebut Ty2, dan empattruncated LTR elements disebut delta sequences. Kelima genome-wide repeatsmembentuk 13.5% dari segmen 50-kb, tetapi gambaran ini tidak secara keseluruhan khaspada genom yeast secara keseluruhan. Ketika ke-16 kromosom yeast dipertimbangkan, jumlah total sekuens yang diambil oleh genome-wide repeats hanya 3.4% dari total. Padamanusia, genome-wide repeats membentuk 44% genom. Gambaran yang muncul adalah bahwa organisasi genetik pada genom yeast lebih ekonomisdibandingkan pada manusia. Gen-gen lebih kompak/padat, memiliki lebih sedikit intron dan ruang antara gen relative pendek, dengan jauh lebih sedikit ruang yang diambil oleh genome-wide repeats dan sekuens-sekuens non-coding. Hipotesis bahwa organism yang lebih komples mengandung genom yang kurang kompak jugaterdapat pada spesies-spesies lain yang diteliti. Gambar diatas juga menunjukkan segmen 50 kbgenom dari lalat buah. Jika kita sependapat bahwa lalat buah lebih kompleks daripada sel yeasttetapi kurang kompleks dibandingkan genom manusia, maka kita akan menduga bahwaorganisasi genom lalat buah akan berada di antara yeast dan manusia.
- 5. Pada gambar, segmen 50kb genom lalat buah memiliki 11 gen, lebih dari gen pada segmen manusia, tetapi kurang darigen pada lalat buah. Semua gen ini discontinuous. Hal ini sesuai ketika keseluruhan sekuensgenom dari 3 organisme dibandingkan (Tabel 2). Densitas gen pada genom lalat buah adalahintermediet antara densitas genom pada yeast dan manusia. Rata-rata gen lalat buah memilikilebih banyak intron daripada rata-rata gen yeast tetapi tetap tiga kali lebih sedikit dibandingkanrata-rata gen manusia.Tabel 2. Kekompakan genom yeast, lalat buah dan manusia (Brown, 2002 ) Karakteristik Yeast Lalat buah Manusia Gene density (average number per Mb) 479 76 11 Introns per gene (average) 0.04 3 9 Amount of the genome that is taken upby 3.40% 12% 44% genome-wide repeats Transkripsi pada Eukariot Struktur gen Secara umum hampir sama ama prokariot ada promotor, bagian struktural dan terminator. Yg beda pada bagian strukturalnya Bagian struktural pada eukariot Nah kenapa bagian struktural/ coding regionnya beda? karena kalo di eukariot ada bagian intron dan ekson. Intro (intervening sequences) merupakan sekuens yg tidak mengkode asam amino. Kalo di gambar yg warnanya biru mudaagak ijo. Ntr bagian ini akan dibuang saat pematangan RNA. Ekson sekuens yg nantinya dikode jd asam amino. kalo di gambar warnanya merah
- 6. Mekanisme Transkripsi Kalo di eukariot RNA polimerasenya beda-beda ada RNA polimerase I, II dan III. Ntr penggunaannya dalam sintesis molekul beda. Sebelum RNA polimerase nempel di promotor, ada faktor transkripsi yang bantu ng-guide si RNA polimerase. Kalo RNA polimerase I guidenya SL1 dan UBF, RNA polimerase II dibantu ama TFIIA, TFIIB, TFIID, TFIIE, TFIIF, TFIIH dan TFIIJ (banyak banget ==”). kalo RNA polimerase III dipandu ama TFIIIA, TFIIIB, TFIIIC ama protein TBP. Nah fyi aja faktor TBP merupakan protein yg diperlukan kalo gen2 g punya TATA box.Trus setelah si RNA polimerase dibantu ama faktor transkripsi (TF) ke TATA box baru terjadi proses elongasi dan berhenti sampeketemu terminator. Proses pasca-transkripsi Ya karena adanya intron pada eukariot, makanya mRNA yg dihasilkan g bisa langsung dikeluarin ke sitosol untuk ditranslasi namunharus diolah dulu. Caranya? 1. Splicing Merupakan proses pembuangan intron dan penyambungan ekson. Awalnya RNA hasil transkripsi pd eukariot disebut pre-mRNA karenamasih ada intronnya. Trus intron akan dipotong dan ekson2 disambung menjadi mRNA matang (mature mRNA). Untuk lebih jelasnya: Intron dipotong pake spliceosome. lalu penyambungan ekson2 pake enzil ligase. 2. Poliadenilasi Merupakan proses penambahan poliA (rantai AMP) pada ujung 3′ nukleotida mRNA. Fungsinya? untuk meningkatkan stabilitas mRNA dan meningkatkan efisiensi translasinya. 3. Capping Penambahan tudung mRNA berupa molekul 7-metilguanosin.
- 7. Fungsinya ada 4: Melindungi mRNA dari degradasi Meningkatkan efisiensi translasi mRNA Meningkatkan pengangkutan mRNA dari nukelus ke sitoplasma Meningkatkan efisiensi proses splicing
- 8. B. Genom prokariot Genom prokariot berbeda dengan genom eukariot. Terdapat beberapa overlap dalam ukuranantara genom prokariotik terbesar dengan prokariotik terkecil. Tetapi secara keseluruhanprokarotik genom berukuran lebih kecil. Misalnya genom E. coli K12 adalah 4639 kb, hanya 2/5dari genom yeast dan hanya memiliki 4405 gen. Organisasi fisik genom juga berbeda antaraeukariot dengan prokariot. Pandangan tradisional adalah bahwa seluruh prokariot memiliki satu molekul DNA sirkular . Selain ‘kromosom’ tunggal ini, prokariot juga dapat memiliki gen tambahan yang independen, sirkular yang disebut plasmid (Gambar 3). Gambar 3. Plasmid adalah DNA sirkular kecil yang terdapat pada sel prokariot. Gen yang dibawa oleh plasmid berguna, karena mengkode sifat-sifat ketahanan terhadapantibiotik atau kemampuan untuk memanfaatkan komponen kompleks seperti toluene sebagaisumber karbon. Tetapi prokariot dapat bertahan secara efektif tanpa plasmid. Prokariotmenunjukkan keragaman dalam organisasi genom. E. coli memiliki genom unipartite, tetapiprokariot lainnya lebih kompleks. Misalnya Borrelia burgdorferi B31, memiliki kromosom linier911 kb, membawa 853 gen, dilengkapi dengan 17 atau 18 molekul linier dan sirkuler, yangkeseluruhannya menyumbangkan 533 kb dan paling tidak 430 gen. Genom multipartite dikenalpada banyak bacteria dan arkaea.Genom prokariotik lebih kompak dibandingkan genom yeast dal eukariot tingkat bawah lainnya.Seperti terlihat pada Gambar 2.2E yang memperlihatkan segmen 50-kb genom E. coli K12.Terlihat bahwa terdapat lebih banyak gen dan kurang ada ruang diantaranya, dengan 43 genmengambil tempat 85.9% segmen. Beberapa gen terlihat tidak memiliki ruang diantaranya, thrAdan thrB, misalnya dipisahkan dengan sebuah nukleotida tunggal, dan thrC mulai pada nukeotidasegera sesudah nukleotida terakhir pada thrB. Ketiga gen ini adalah contoh dari operon, sebuahkelompok gen yang terlibat dalam sebuah lintasan biokimia (dalam hal ini sintesis asam aminothreonine) dan diekspresikan bersama-sama dengan yang lainnya. Operon
- 9. digunakan sebagaimodel untuk memahami bagaimana ekspresi gen diatur. Secara umum, gen prokariot lebihpendek dibandingkan eukariot, rata-rata panjang sebauh gen bakteri berkisar 2/3 gen eukariot,bahkan setelah intron dihilangkan dari eukariot. Gen bakteri sedikit lebih panjang dibandingkangen arkaea.Dua karakteristik genom prokariot yang dapat dilihat dari Gambar 2.2E adalah, pertama, tidak ada intron pada gen pada segmen dari genom E. coli ini. Bahkan E. coli tidak memiliki gendiscontinuous. Karakteristik kedua adalah infrequency of repetitive sequences. Genom prokariottidak memiliki apapun yang ekivalen terhadap high-copy-number genome-wide repeat familiesyang ditemukan pada genom eukariot. Mereka memiliki sekuen tertentu yang mungkin berulang di dalam genom. Contohnya adalah insertion sequences IS1 dan IS186yang dapat dilihat padasegmen 50-kb pada Gambar 2.2E. Terdapat contoh transposable elements, yaitu sekuen yangdapat berpindah sekeliling genom. Posisi elemen IS1dan IS186 yang ditunjukkan pada Gambar2.2E merujuk pada isolate E. coli tertentu. Jika isolate berbeda yang diperiksa, maka sekuen ISdapat berbeda posisi atau dapat pula absen dari genom. Sebagian besar genom prokariot lainnyamemiliki sangat sedikit sekuen berulang/repeat sequences – secara virtual tidak terdapat padagenom 1.64 Mb dari Campylobacter jejuni NCTC11168 – tetapi terdapat perkecualian, padabakteri meningitis Neisseria meningitidis Z2491, yang memiliki lebih dari 3700 copi dari15 tipeberbeda repeat sequence, secara kolektif membentuk hamper 11% dari genom 2.18 Mb. Transkripsi pada Prokariot Salah satu ciri dari prokariot adalah adanya struktur operon. Operon adalah organisasi daribeberapa gen yang ekspresinya dikendalikan oleh satu promotor. Misal operon lac,padametabolisme laktosa pada bakteri E.coli. Pada waktu ditranskripsi operon lac akan menghasilkan satu mRNA yang membawa kode2 genetik untuk polipeptida berbeda yang disebut dengan mRNA polisistronik.
- 10. Sesuai gambar di atas, pada operon lac punya 3 gen struktural yaitu lac Z, lac Y dan lac A.Masing-masing dari gen itu punya start codon dan stop codon sendiri2 namun ekspresinya tetep dikendaliin ama operon yg sama. Trus waktu ditranskripsi hasilnya 1 mRNA yg bawa kodon-kodon untuk 3 macam polipeptida yang beda. Trus translasinya? nanti akan jd 3 polipeptida yang independen
- 11. Struktur Gen Prokariot Pada prokariot gennya secara umum tersusun atas promotor, bagian struktural, dan terminator Promotor Promotor adalah urutan DNA spesifik yang berperan dalam mengendalaikan transkripsi genstruktural dan terletak di daerah upstream (hulu) dari bagian struktural gen. Fungsi promotor? Sebagai tempat awal pelekatan enzim RNA polimerase yang nantinyamelakukan transkripsi pada bagian struktural Pada prokariot bagian penting promotornya disebut sebagai Pribnow box pada urutan nukleotida-10 dan -35. Biasanya berupa TATA box. Apa fungsi dari pribnow box ? Pribnow box merupakan daerah tempat pembukaan heliks DNAuntuk membentuk kompleks promotor terbuka. Jadi di TATA box itulah DNA dipisahkan dan kalodi luar TATA box helix DNAnya tetep berikatan (beda ama replikasi kan?)
- 12. Operator Operator merupakan urutan nukelotida yang terletak di antara promotor dan bagian strukturaldan merupakan tempat pelekatan protein represor (penekan atau penghambat ekspresi gen). Jikaada represor yang melekat di operator maka RNA polimerase g bisa jalan trus ekspresi gen tidak bisa berlangsung. Kalo di gambar di atas operator disimbolkan dengan warna ungu yg berada di antara promotor(merah) dan structural gene (hijau).Selain adanya supresor ada juga yg namanya enhancer. kalo supresor untuk menghambat nahenhancer kebalikannya, dia malah meningkatkan transkripsi dengan meningkatkan jumlah RNApolimerase. Namun letaknya tidak pada lokasi yg spesifik spt operator, ada yg jauh di upstream atau bahkan downstream dari titik awal transkripsi. Coding Region Gen struktural merupakan bagian yang mengkode urutan nukleotida RNA. Transkripsi dimulaidari sekuens inisiasi transkripsi (ATG) sampai kodon stop (TAA / TGA / TAG).Pada prokariot tidak ada sekuens intron (yg tidak dapat diekspresikan) sehingga semuanyaberupa ekson. Namun kadang pada archaebacteria dan bakteriofag ada yg memiliki intron.
- 13. Terminator Dicirkan dengan struktur jepit rambut / hairpin dan lengkungan yang kaya yang akan urutan GCyang terbentuk pada molekul RNA hasil transkripsi. RNA Polimerase RNA polimerase merupakan enzim yang mengkatalisis proses transkripsi. Kalo susunannyalengkap α2ββ’σ disebut holoenzim. Kalo g ada σ Cuma α2ββ’ disebut core-enzyme.Fungsi subuni2 itu: α = diduga berfungsi dalam penyusunan enzim β = berfungsi dalam pengikatan nukleotida β’ = berfungsi dalam penempelan DNA σ = berfungsi untuk mengarahkan agar RNA polimerase menempel pada promotor. Mekanisme Transkripsi o Inisiasi o Elongasi o Terminasi Daftar Pustaka : Brown, T.A. 2002. Genome. Online book. http://onlinebooks.library.upenn.edu/webbin/book/lookupid?key=olbp36849 Nama : Mutia Rizqa Firdiah NIM : 25010112140145 Kelas : B 2012 Matkul : Biomol