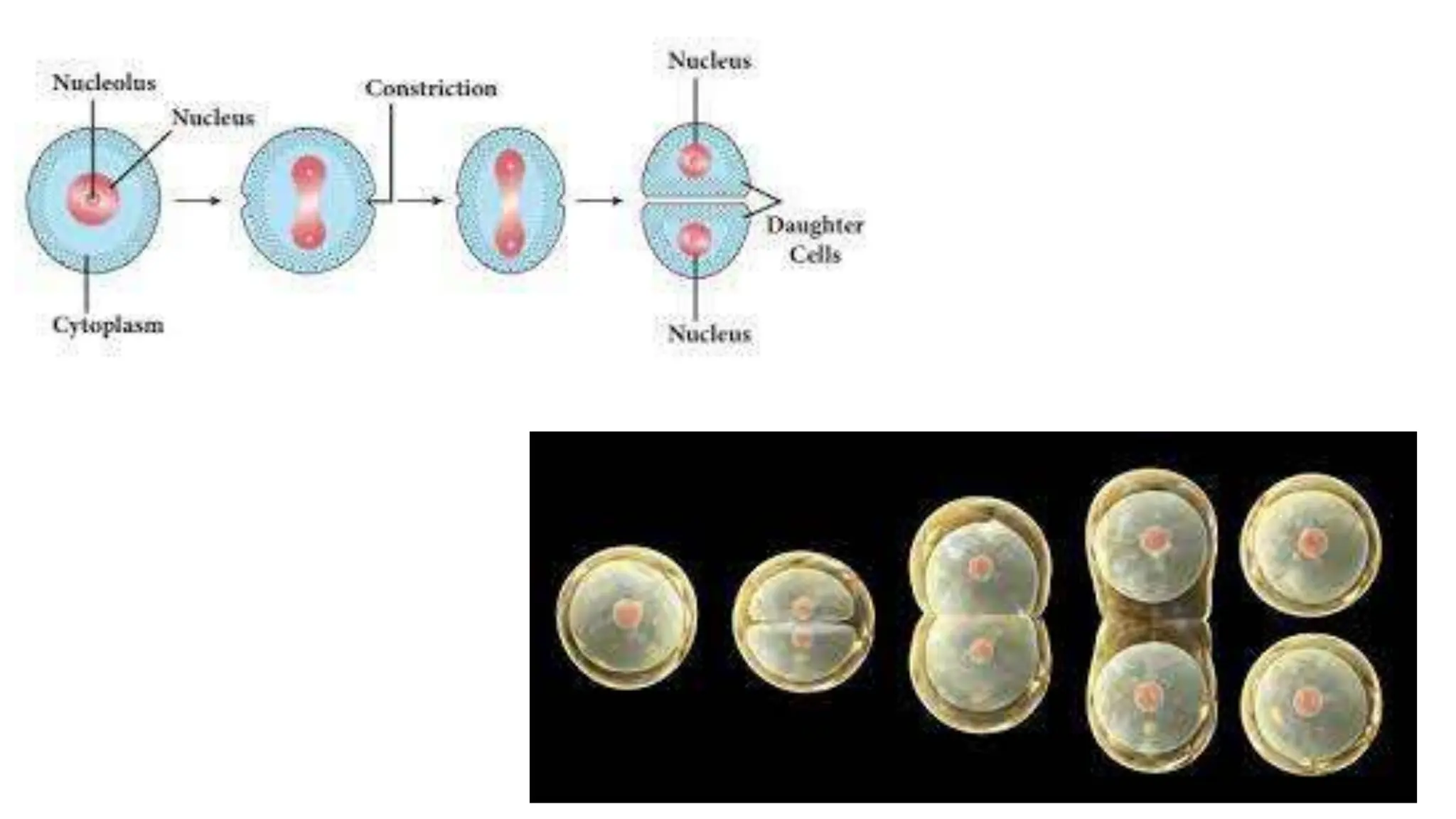
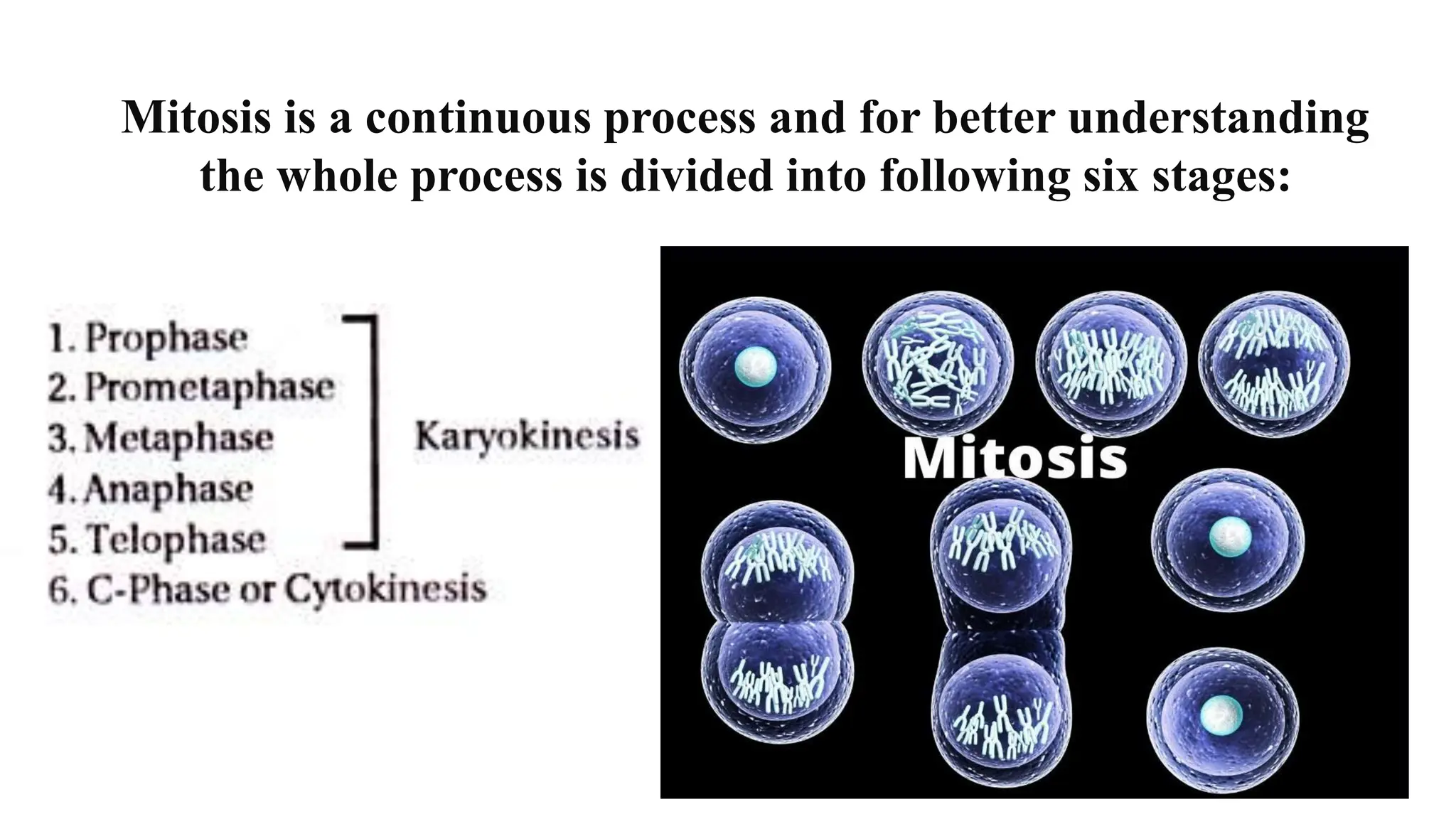
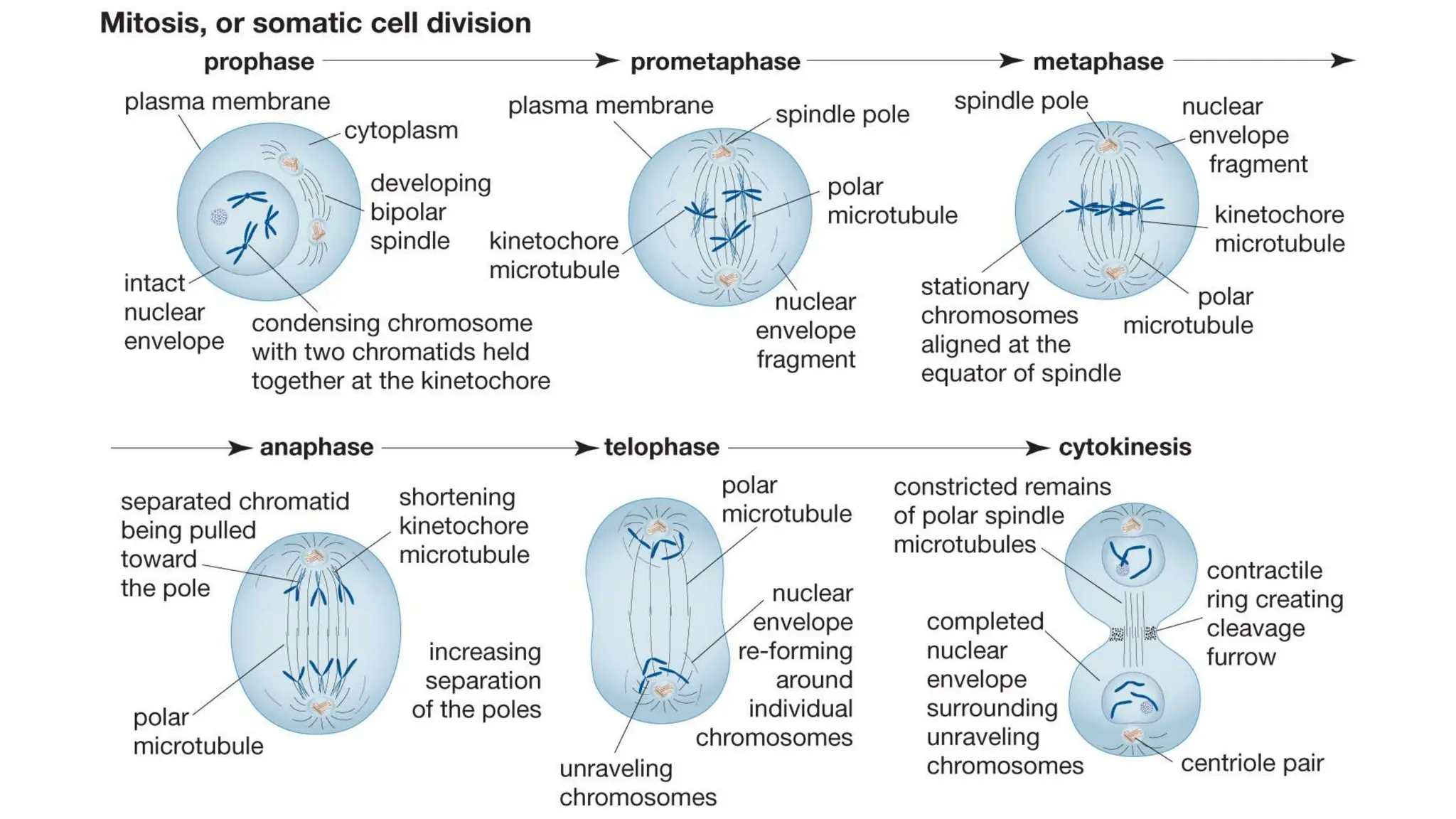
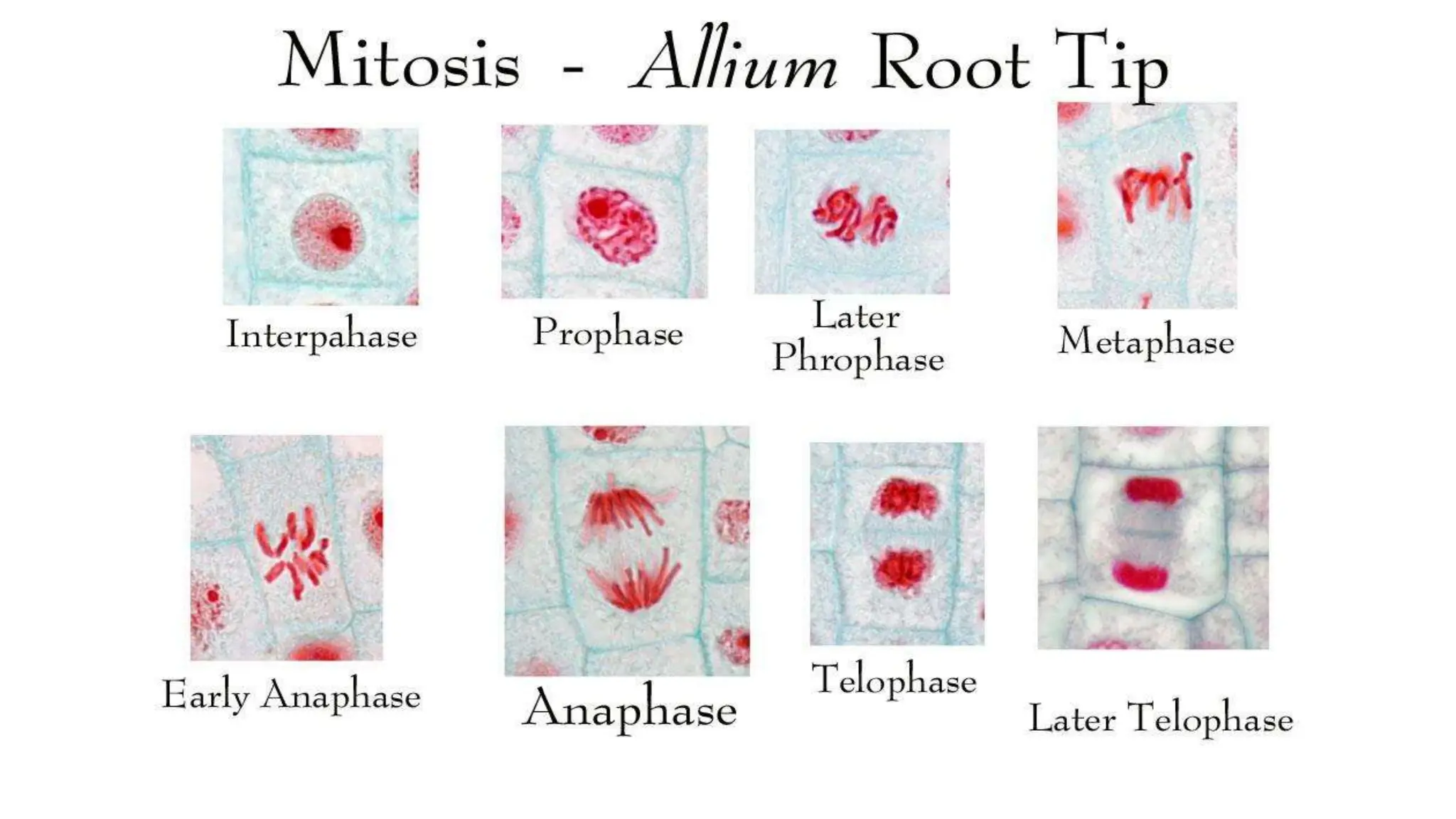
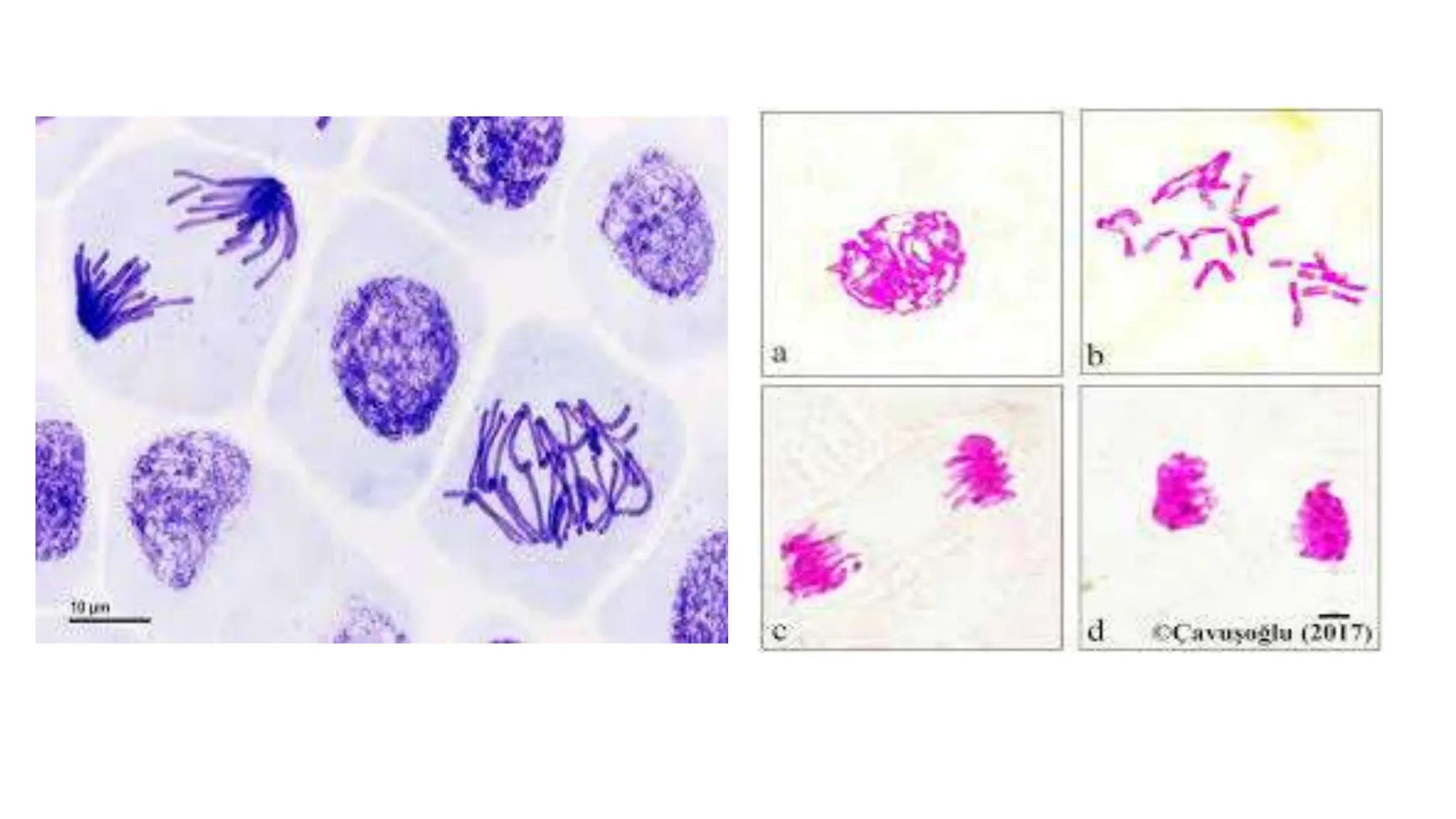
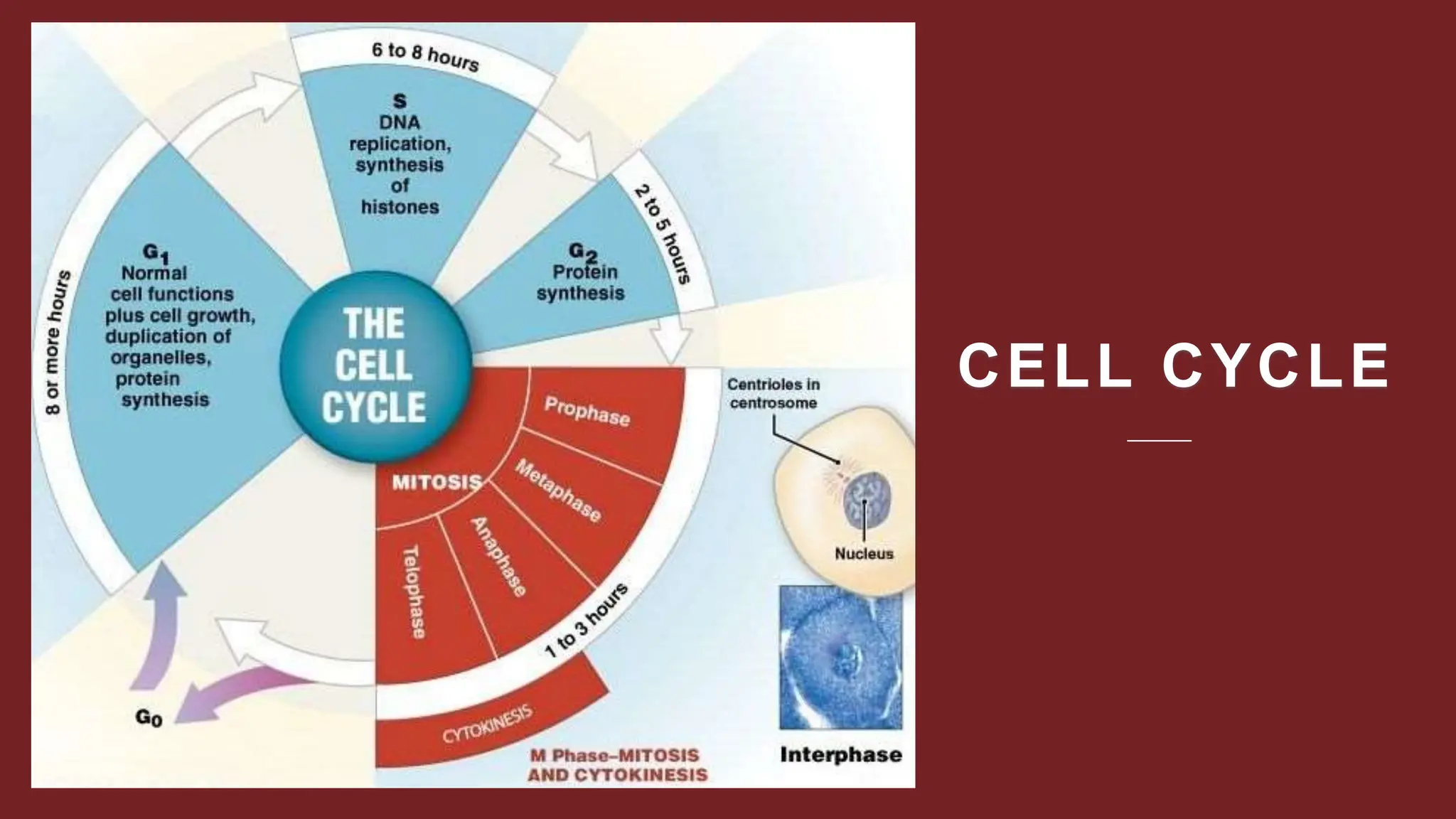
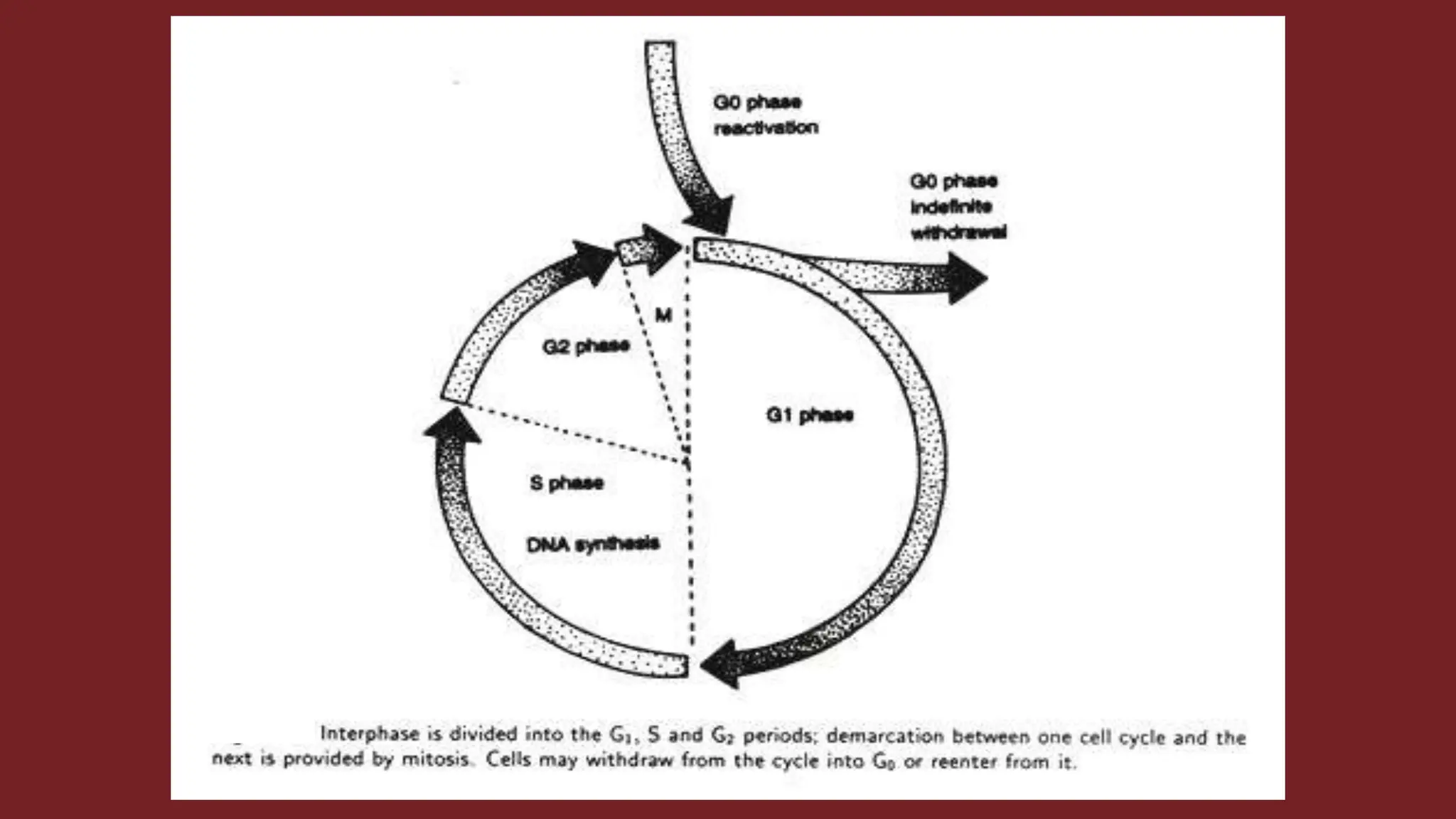
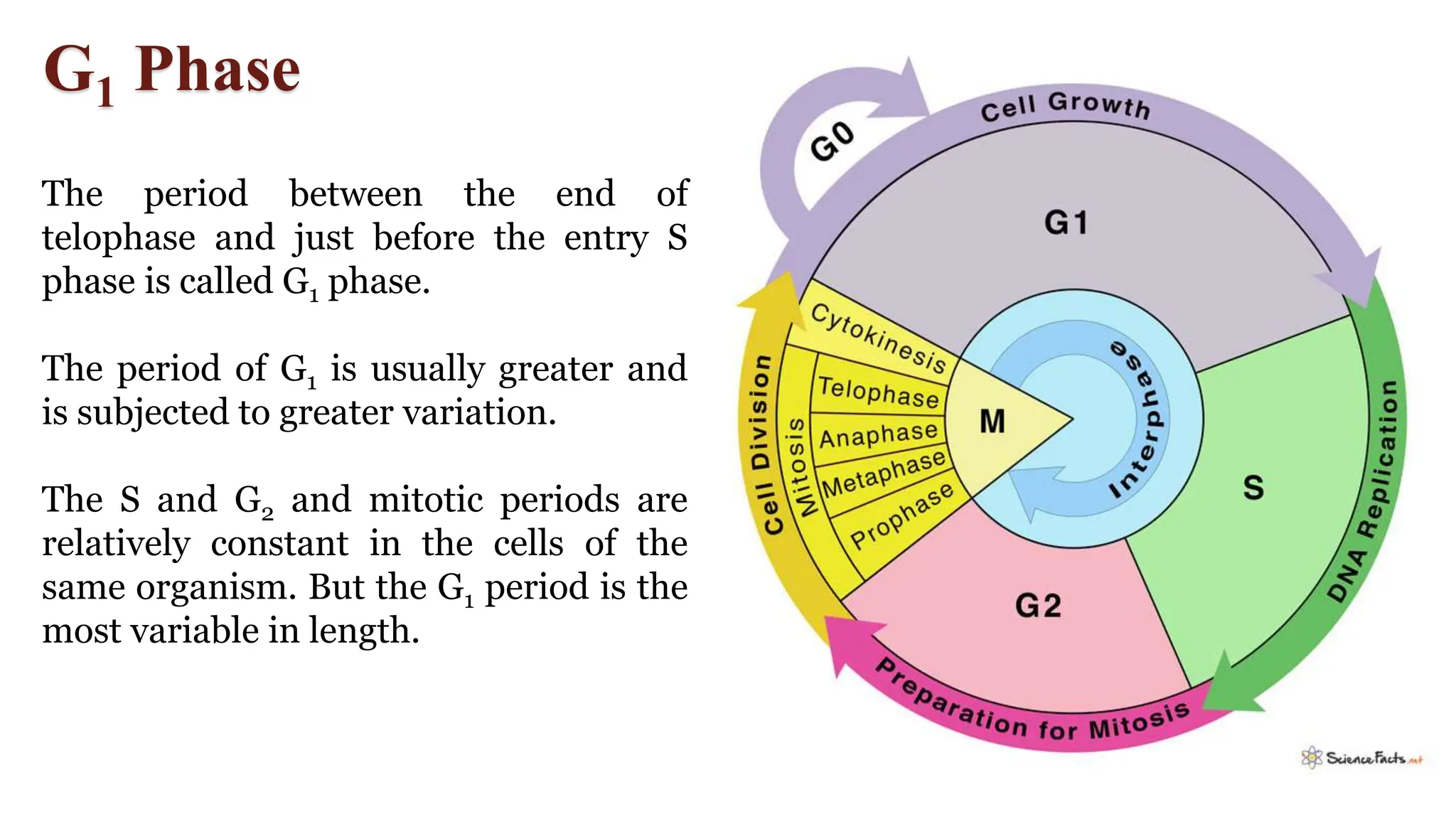
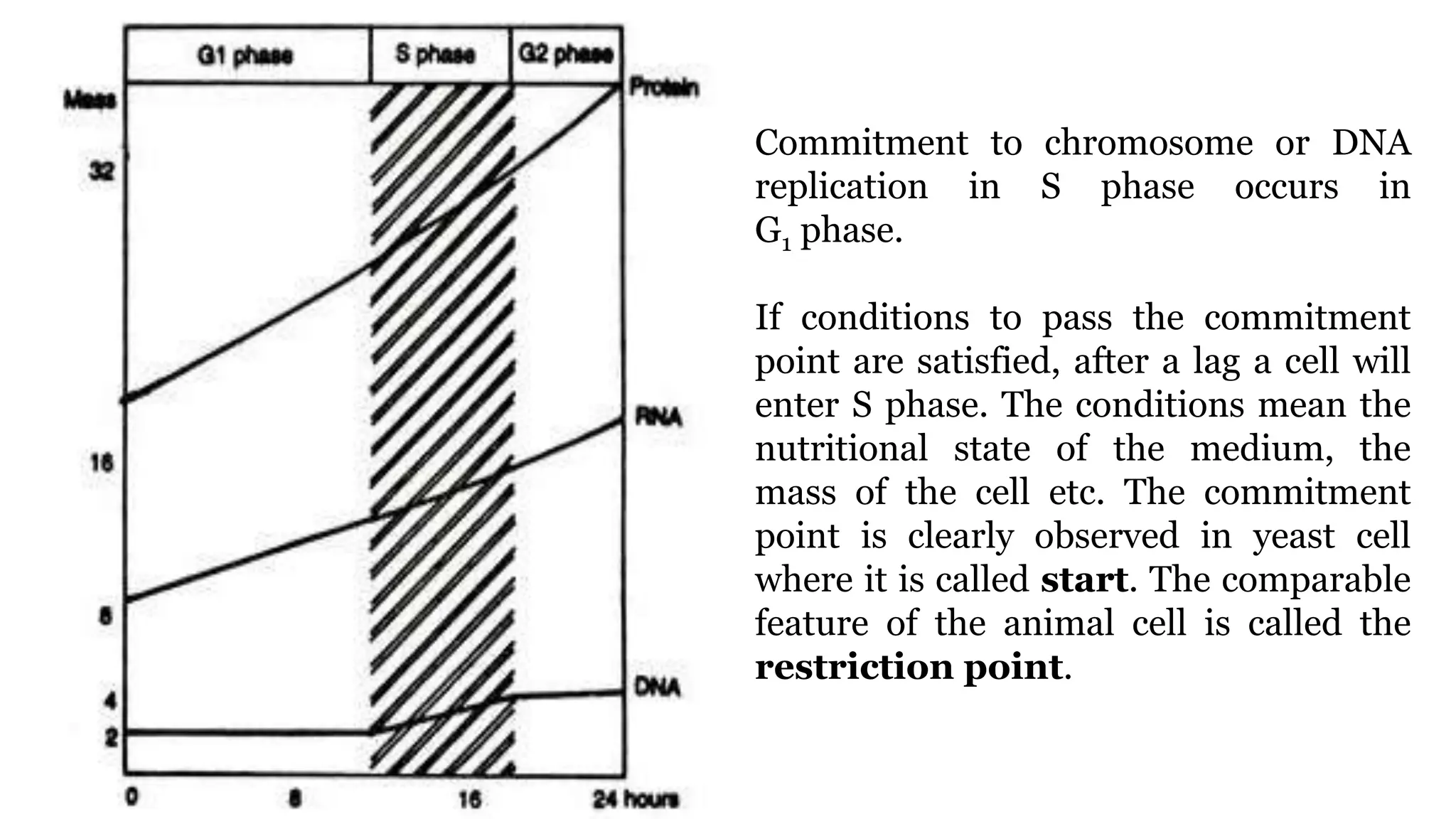
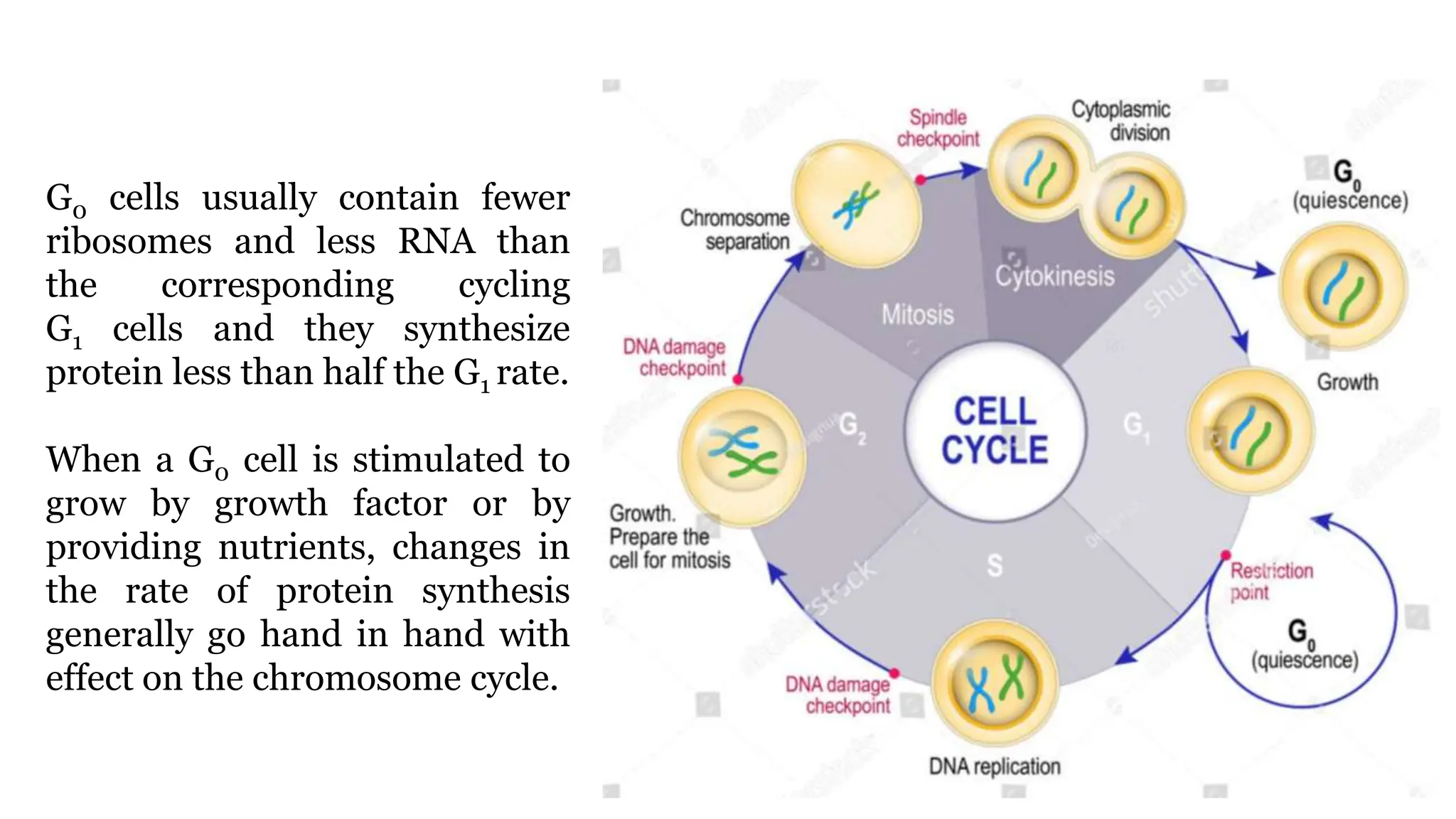
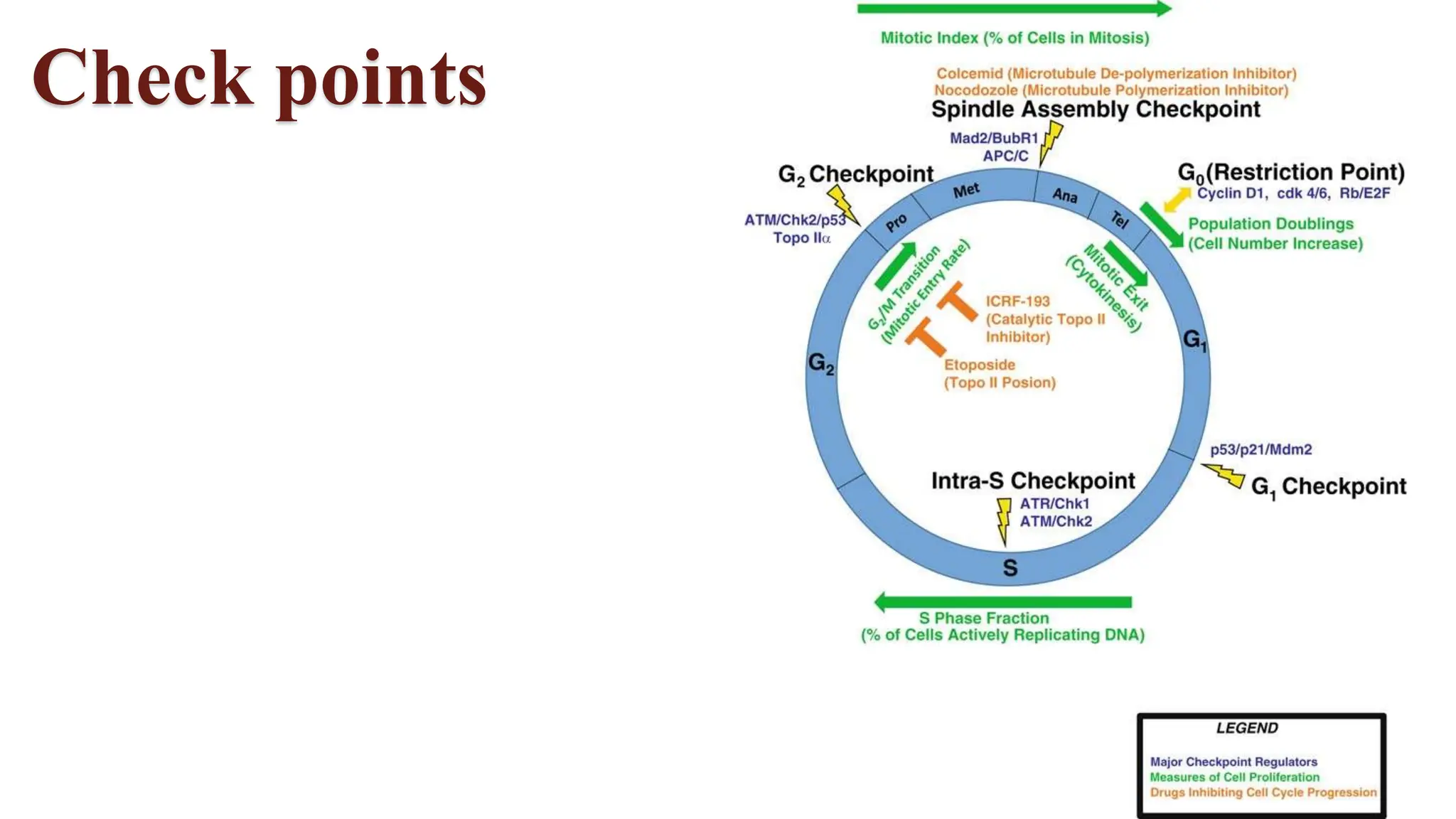
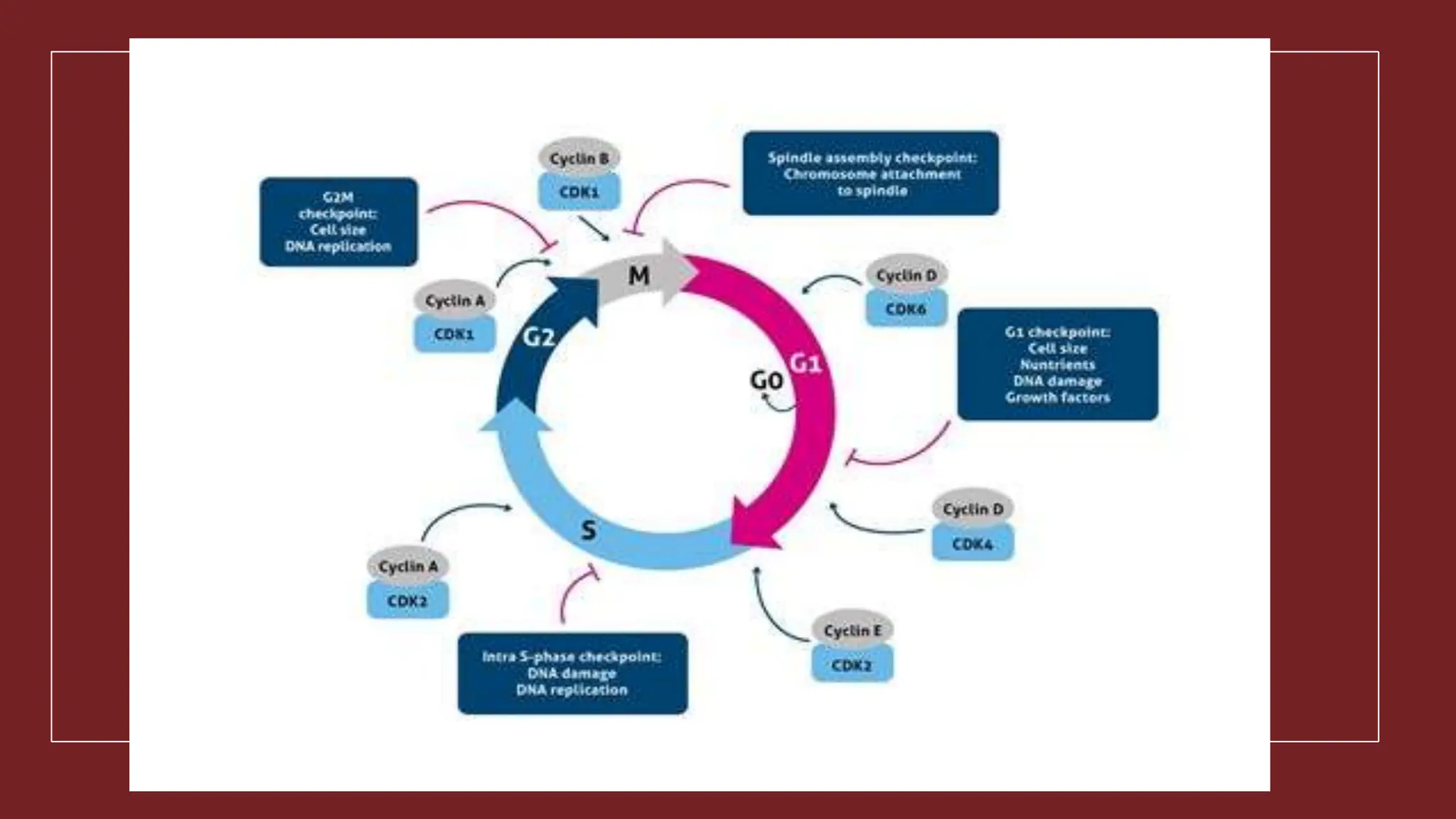
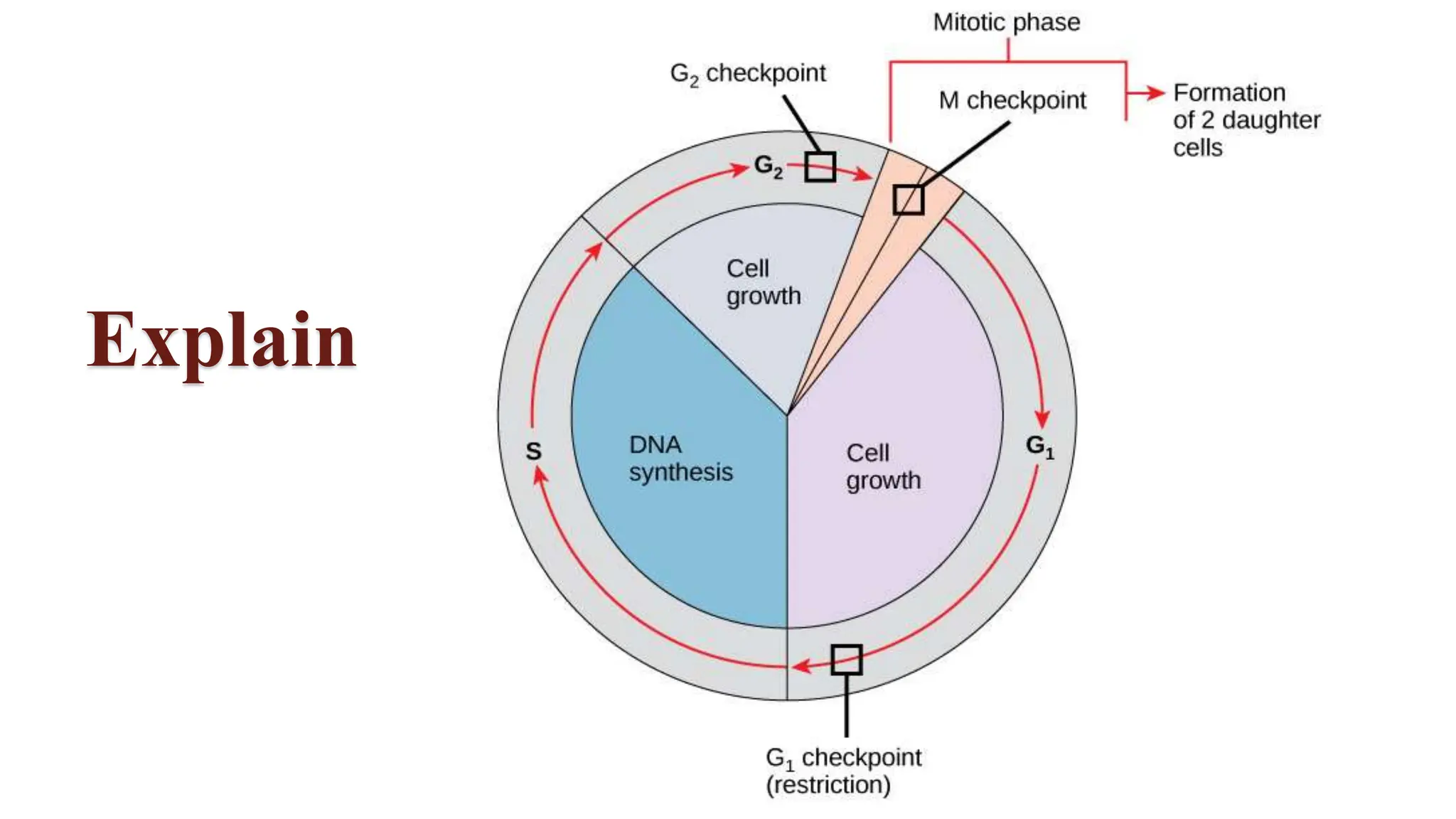
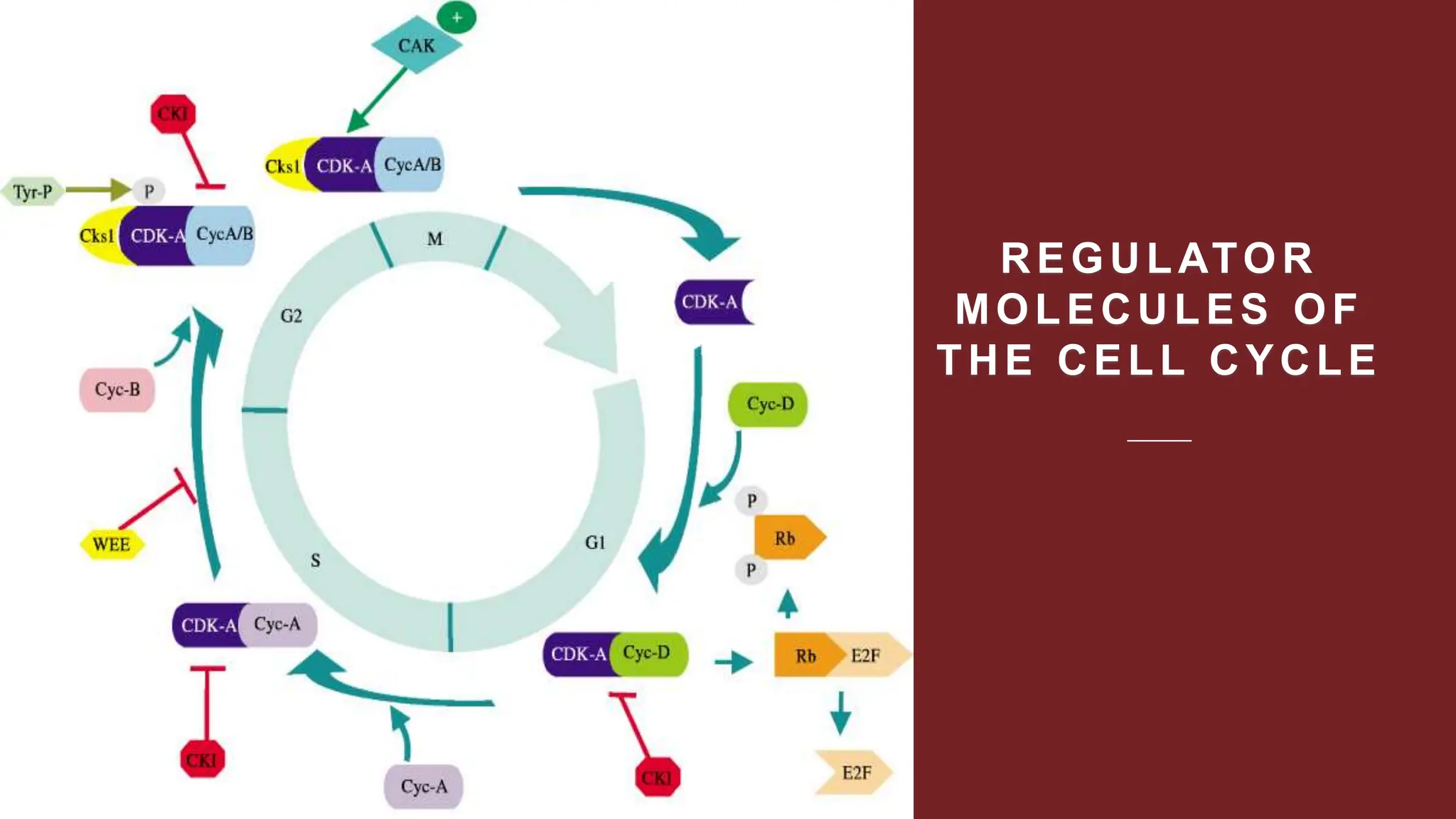
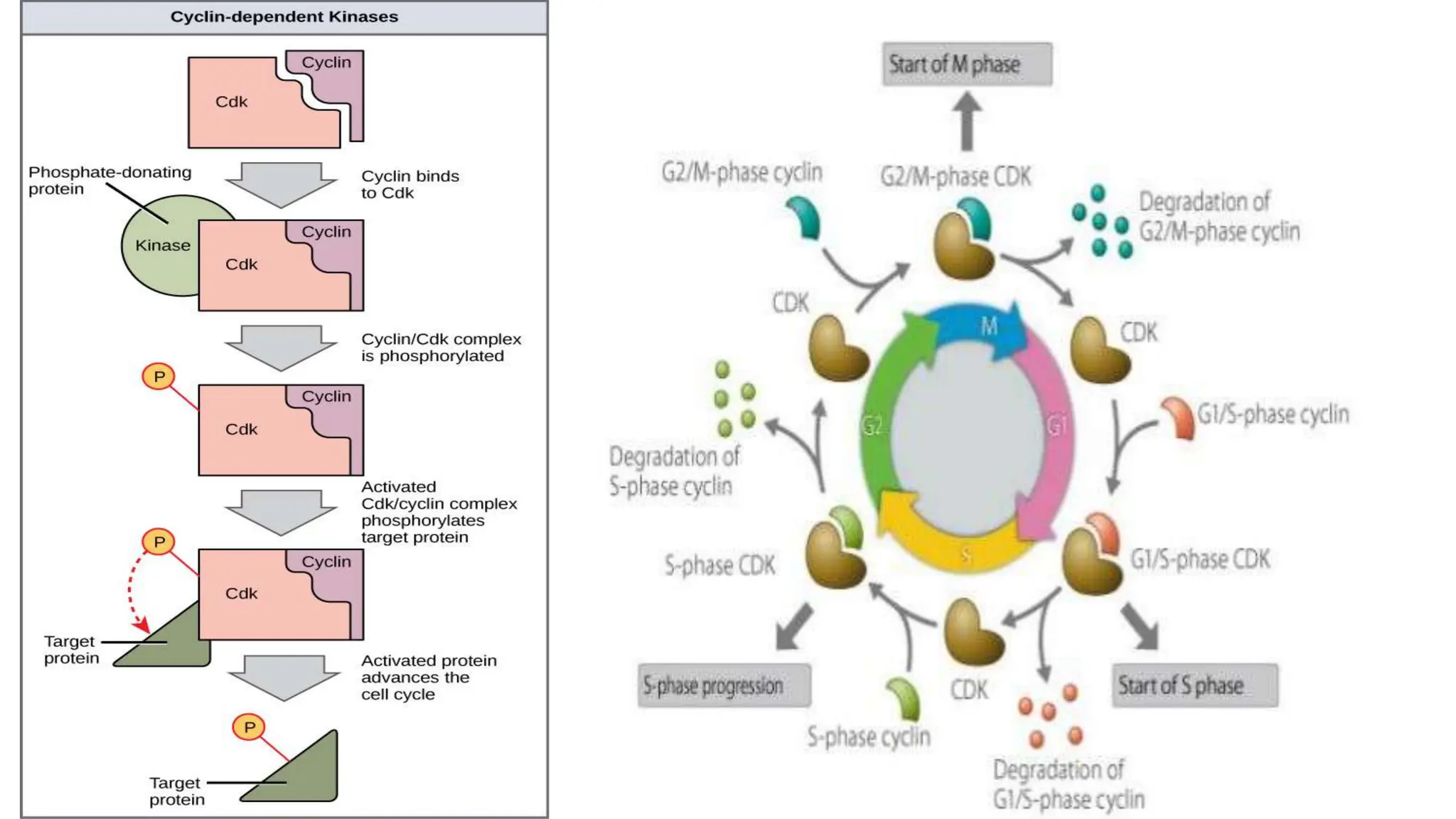
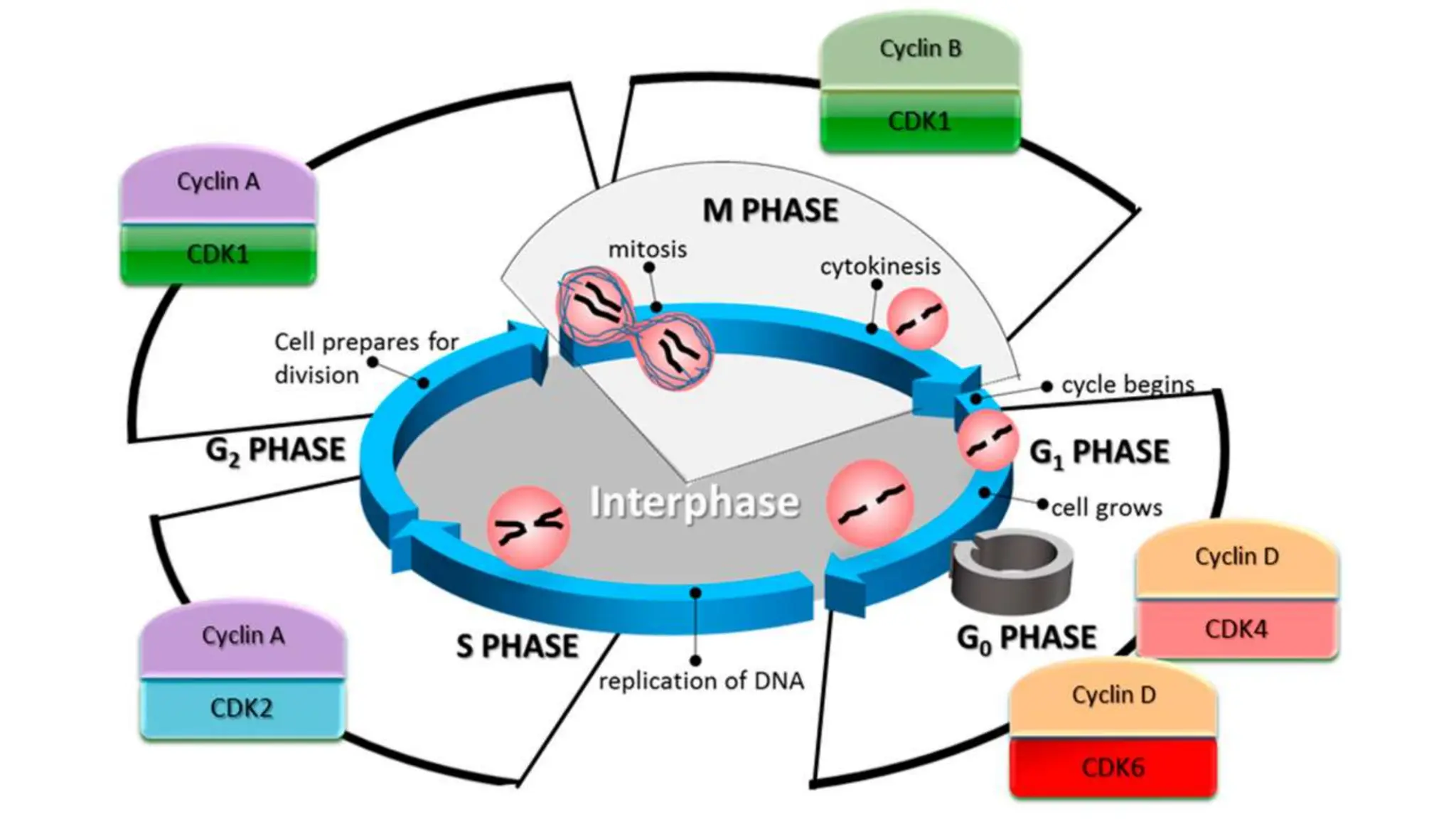
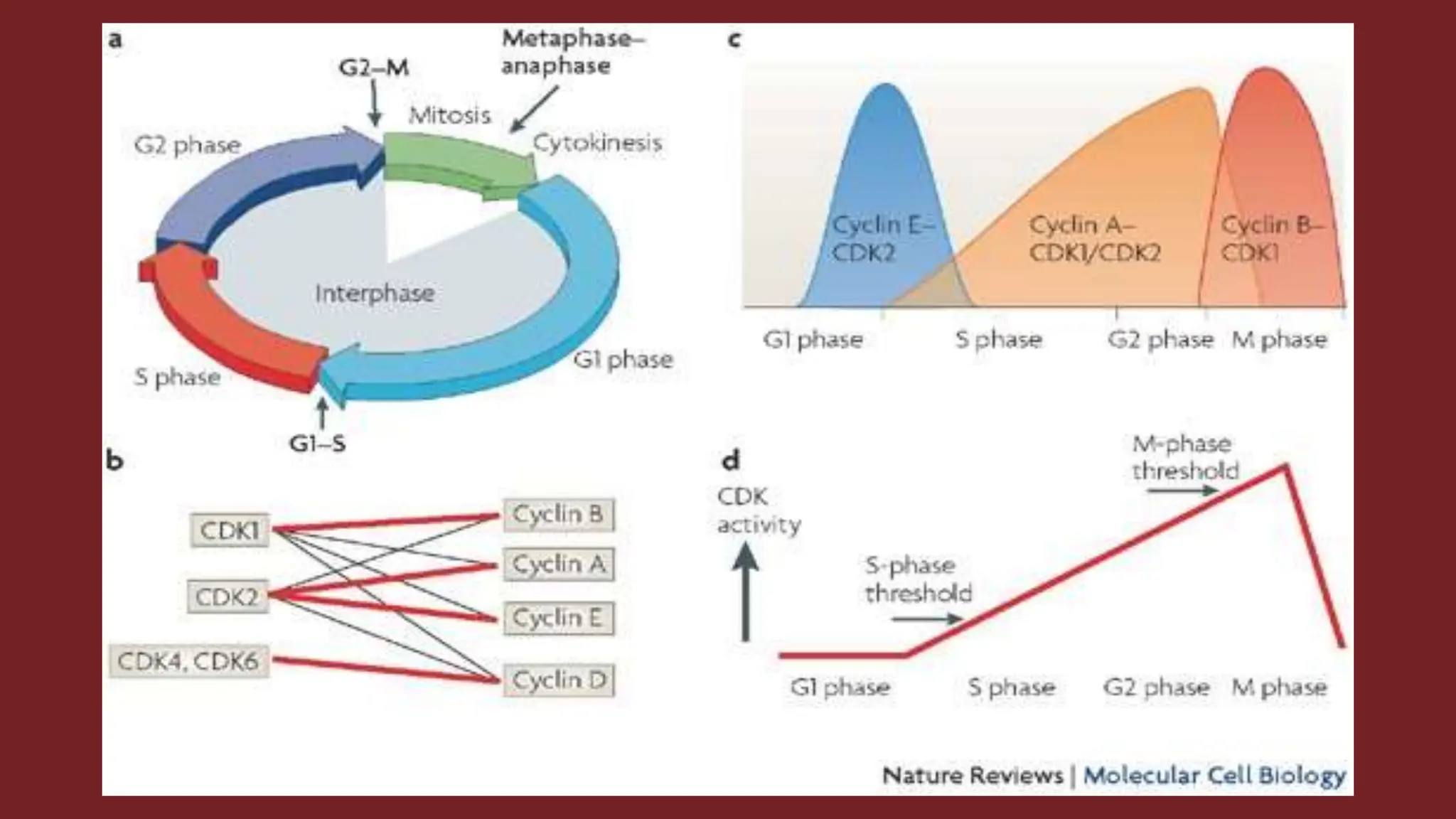
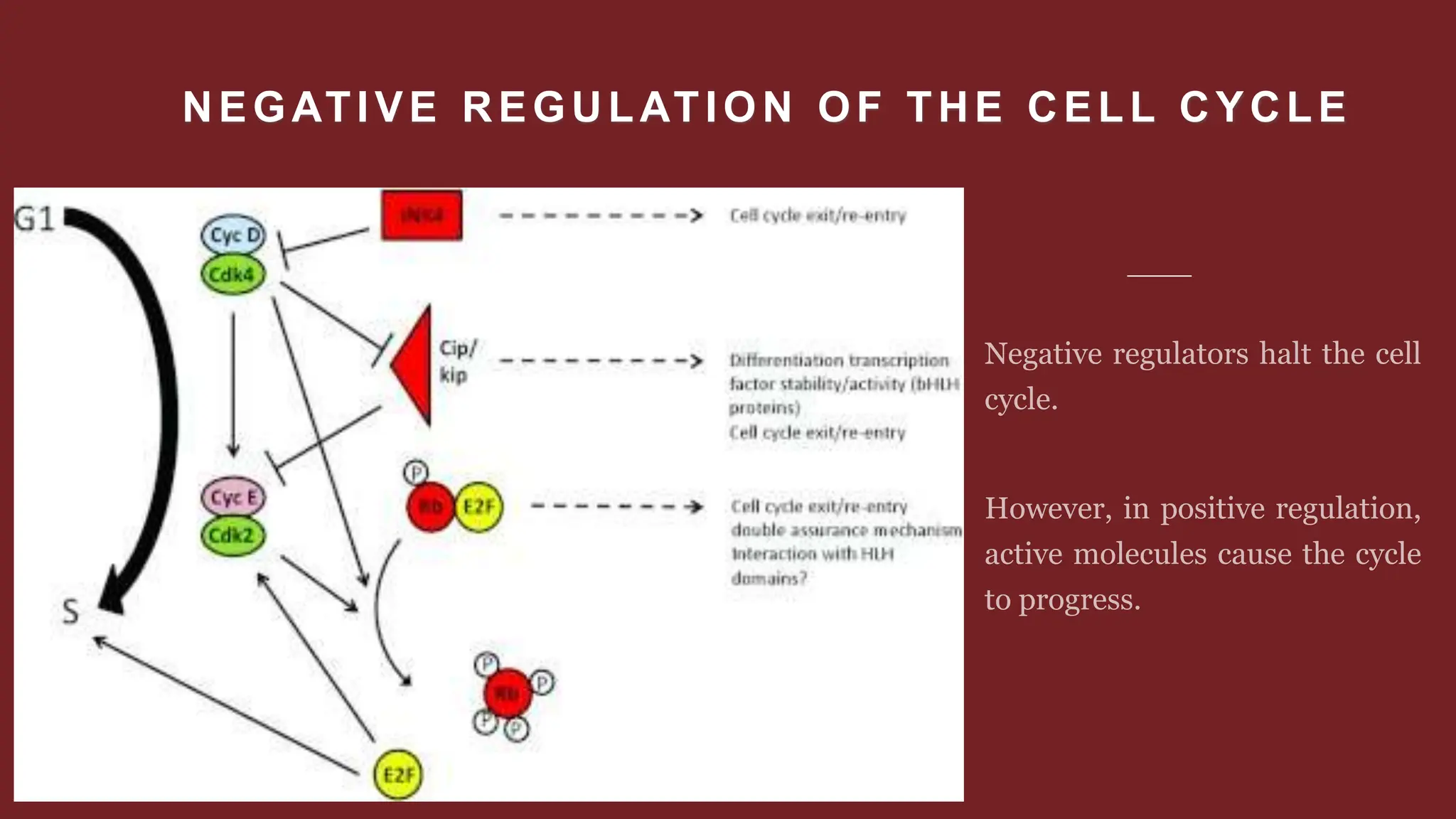
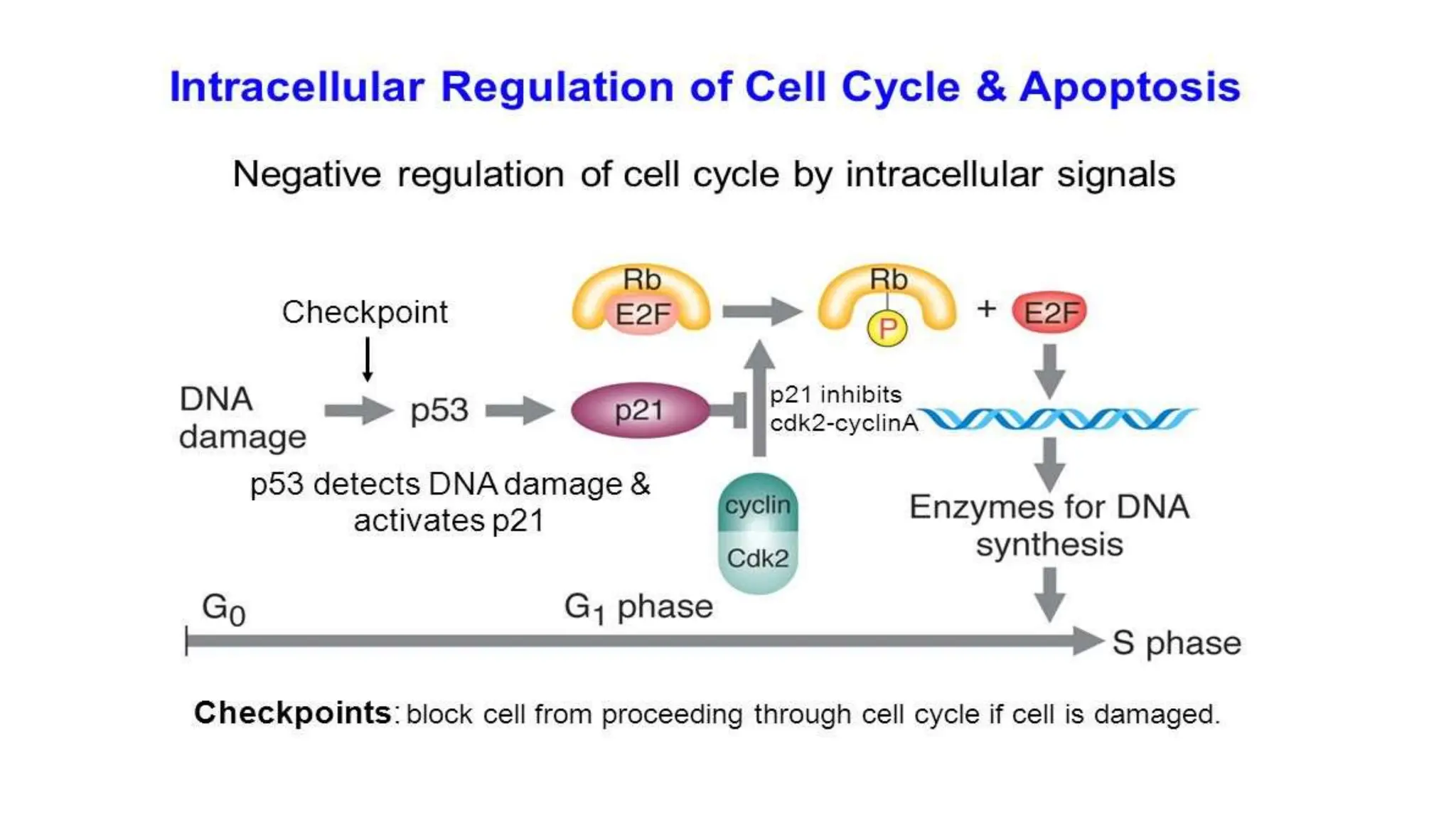
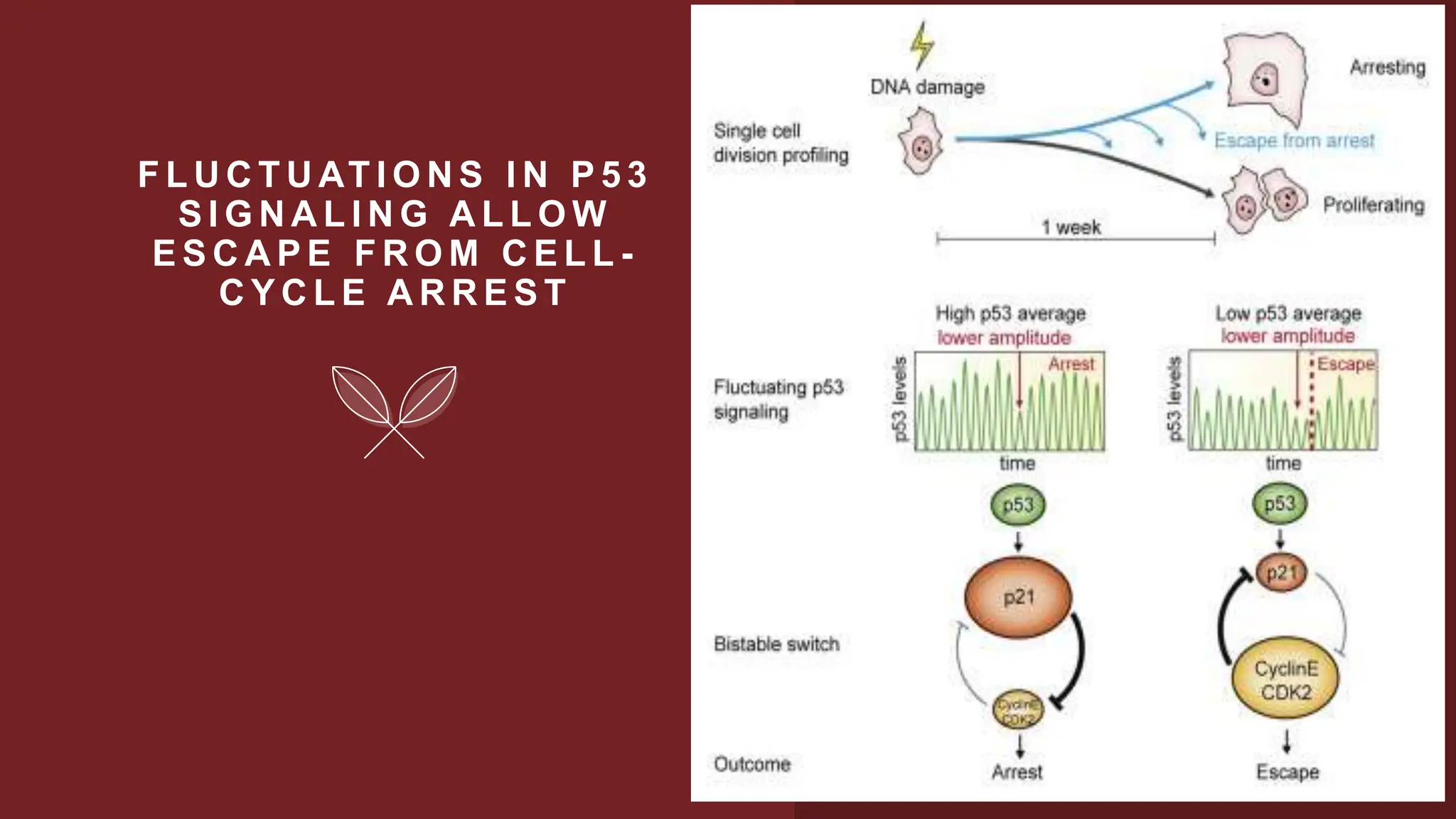
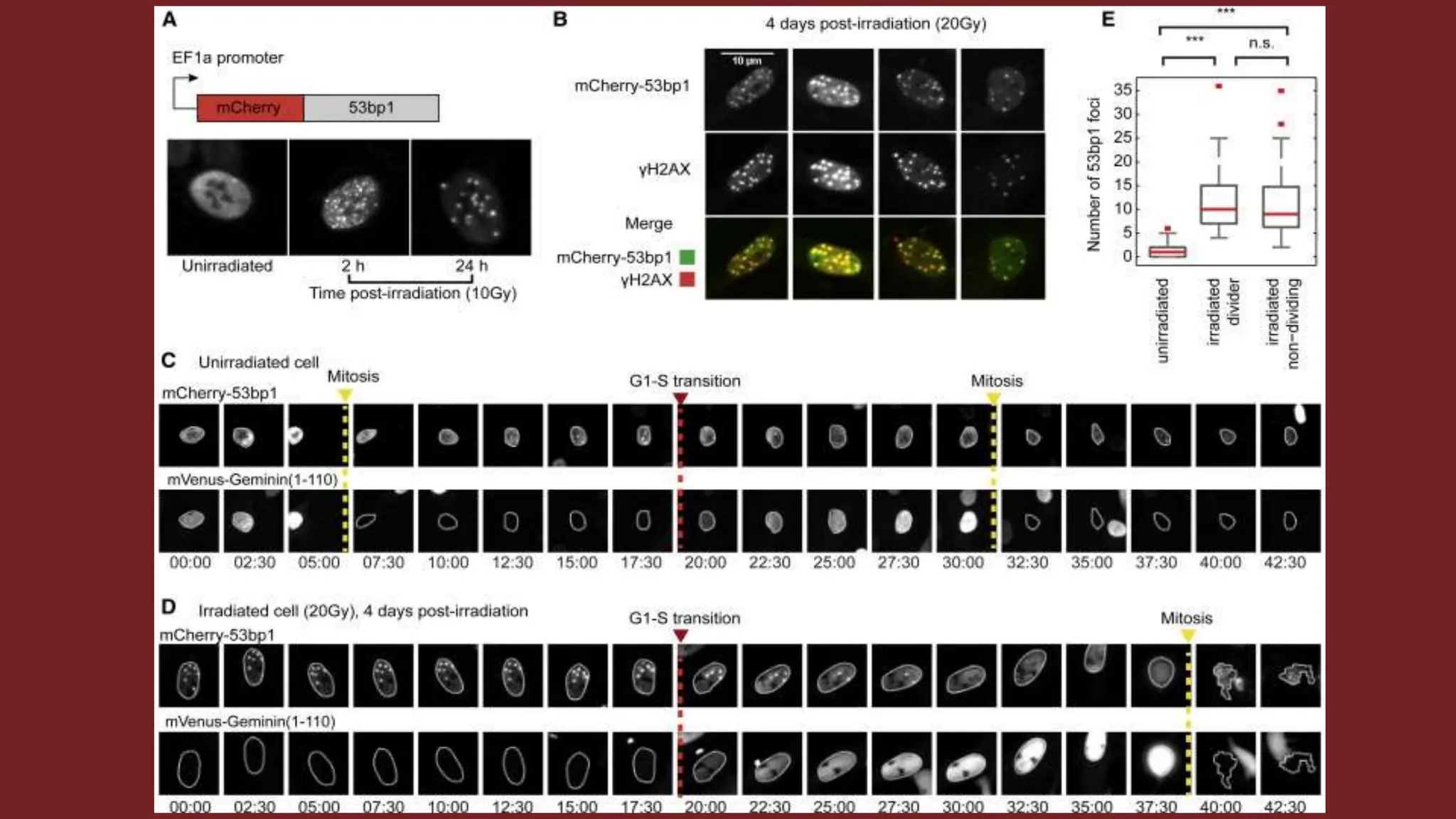
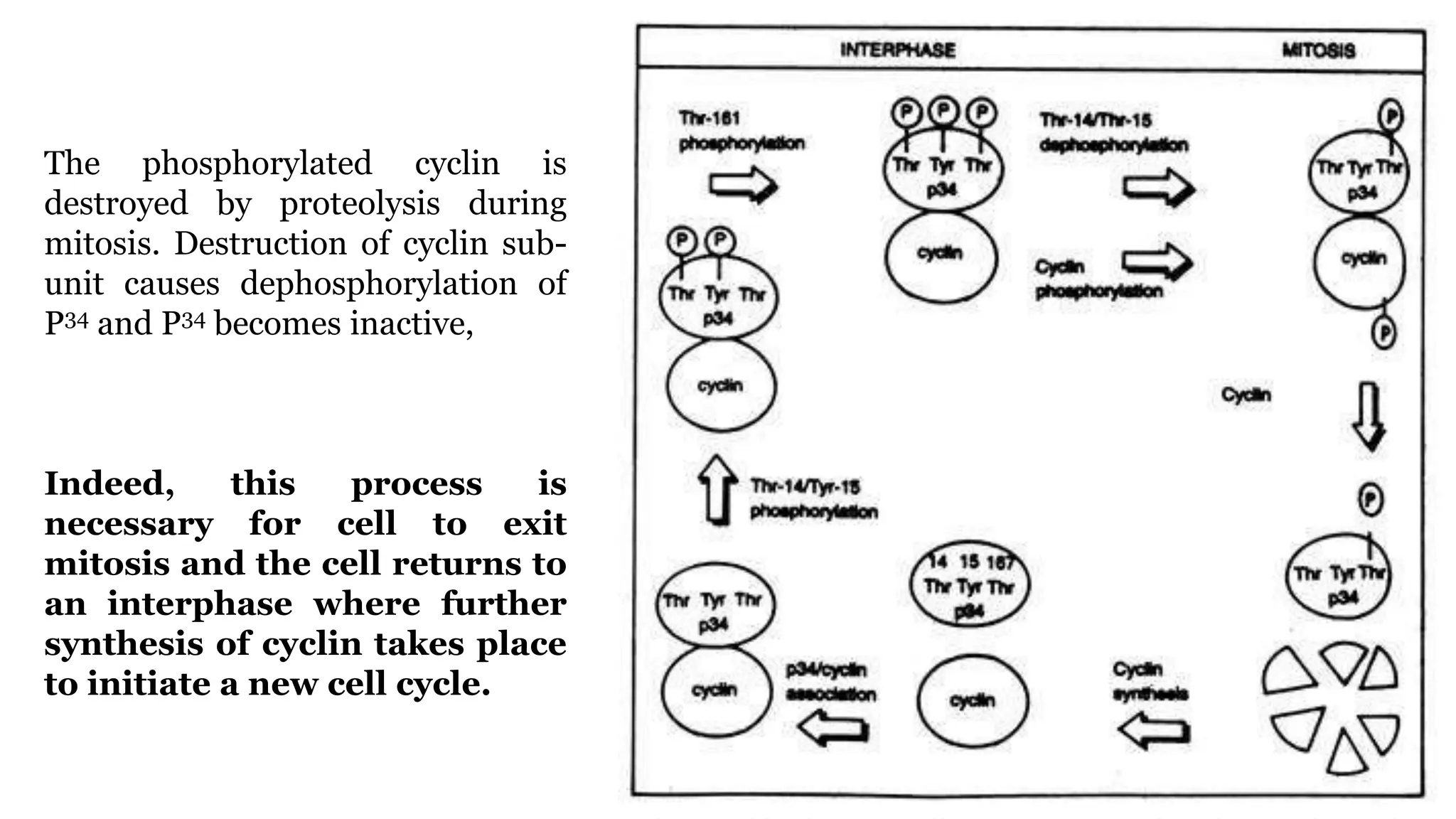
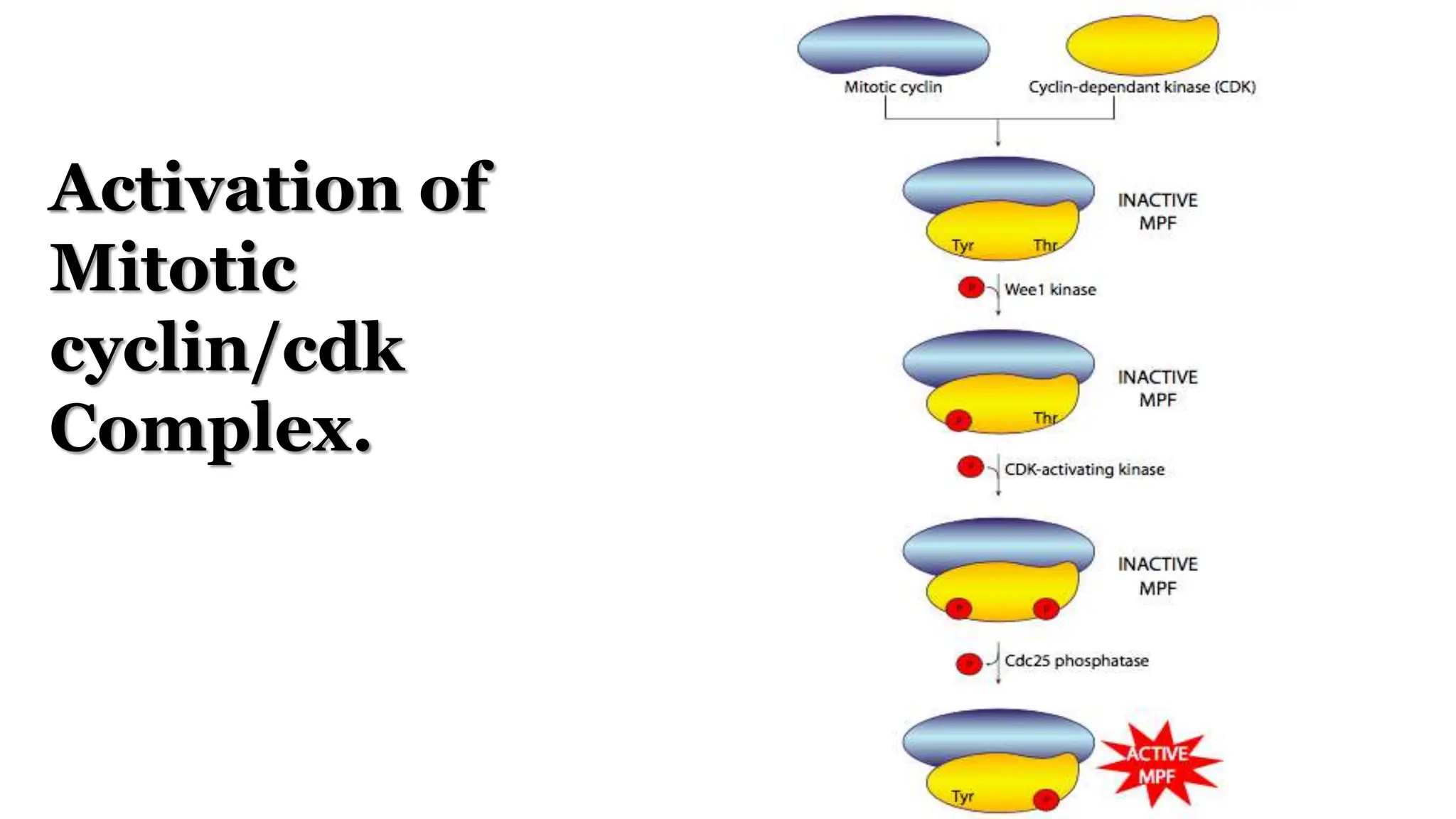
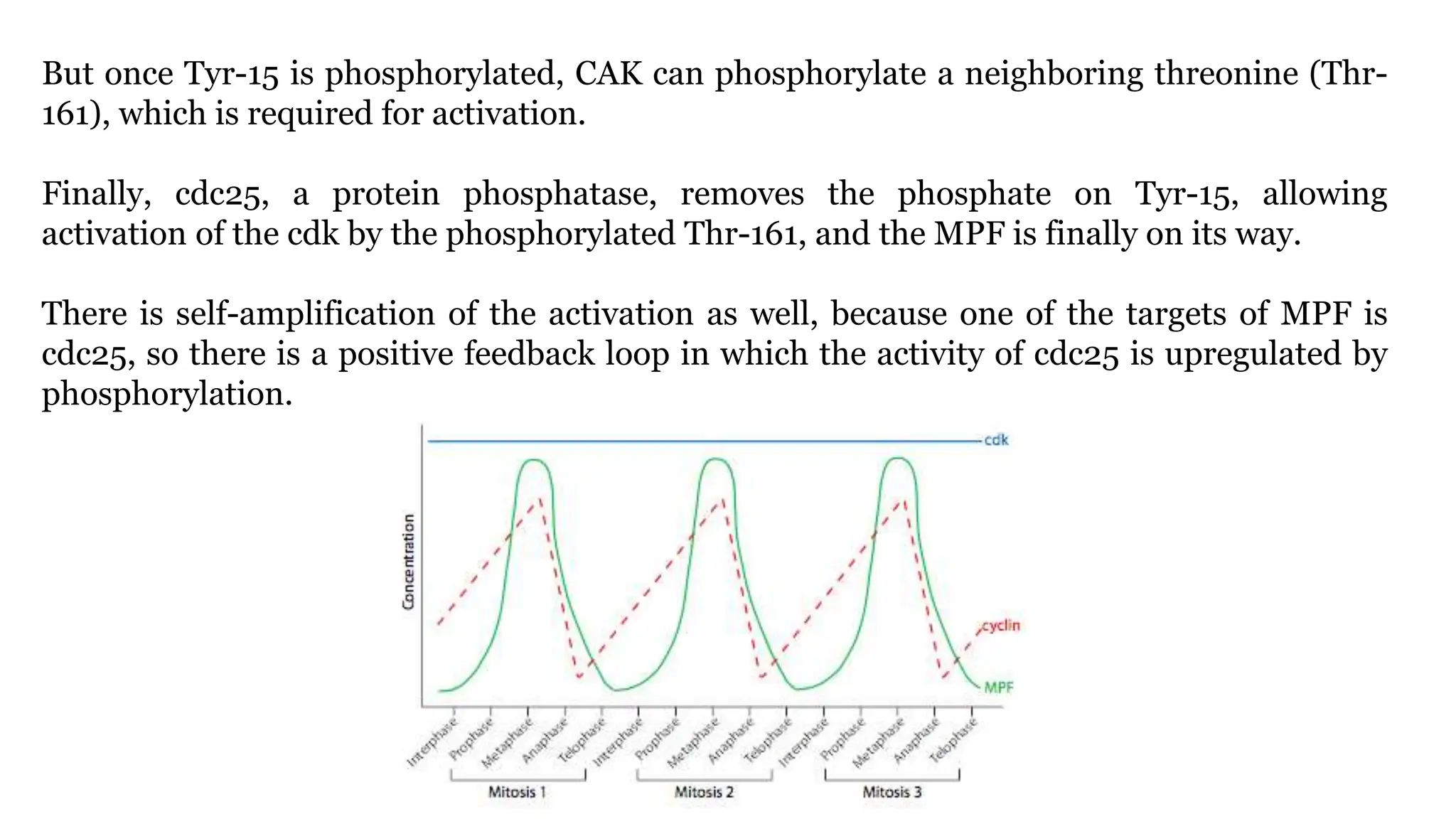
Cell division is essential for life continuity and encompasses processes like asexual reproduction in unicellular organisms and growth and repair in multicellular organisms. The document details the types of cell division (amitosis, mitosis, and meiosis), the phases of the cell cycle (interphase, mitosis, and cytokinesis), and the regulatory mechanisms that control the process, emphasizing the roles of cyclins, cyclin-dependent kinases, and tumor-suppressor proteins. It highlights the importance of checkpoints in the cell cycle to ensure proper cell division and prevent the propagation of damaged DNA.