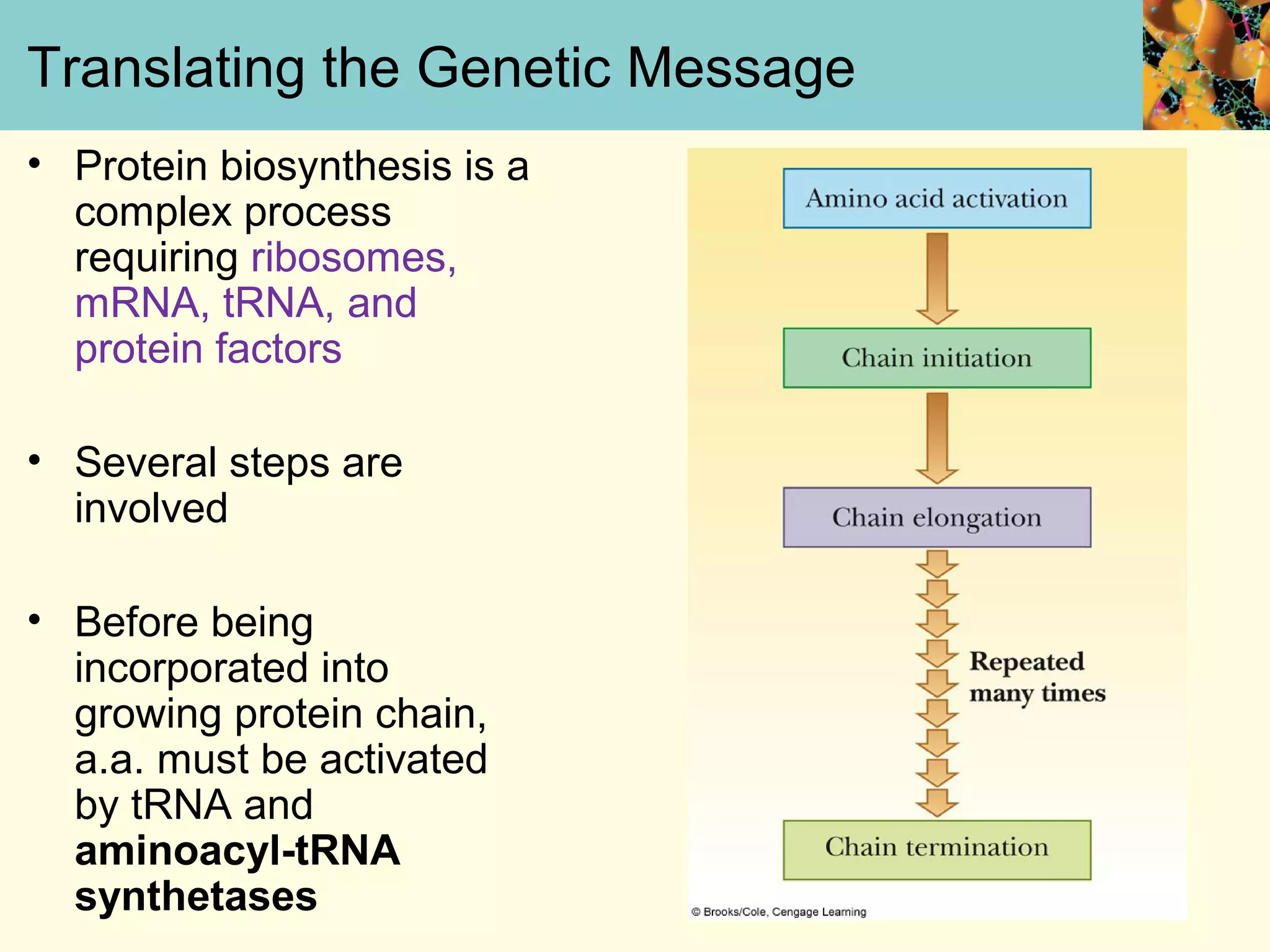
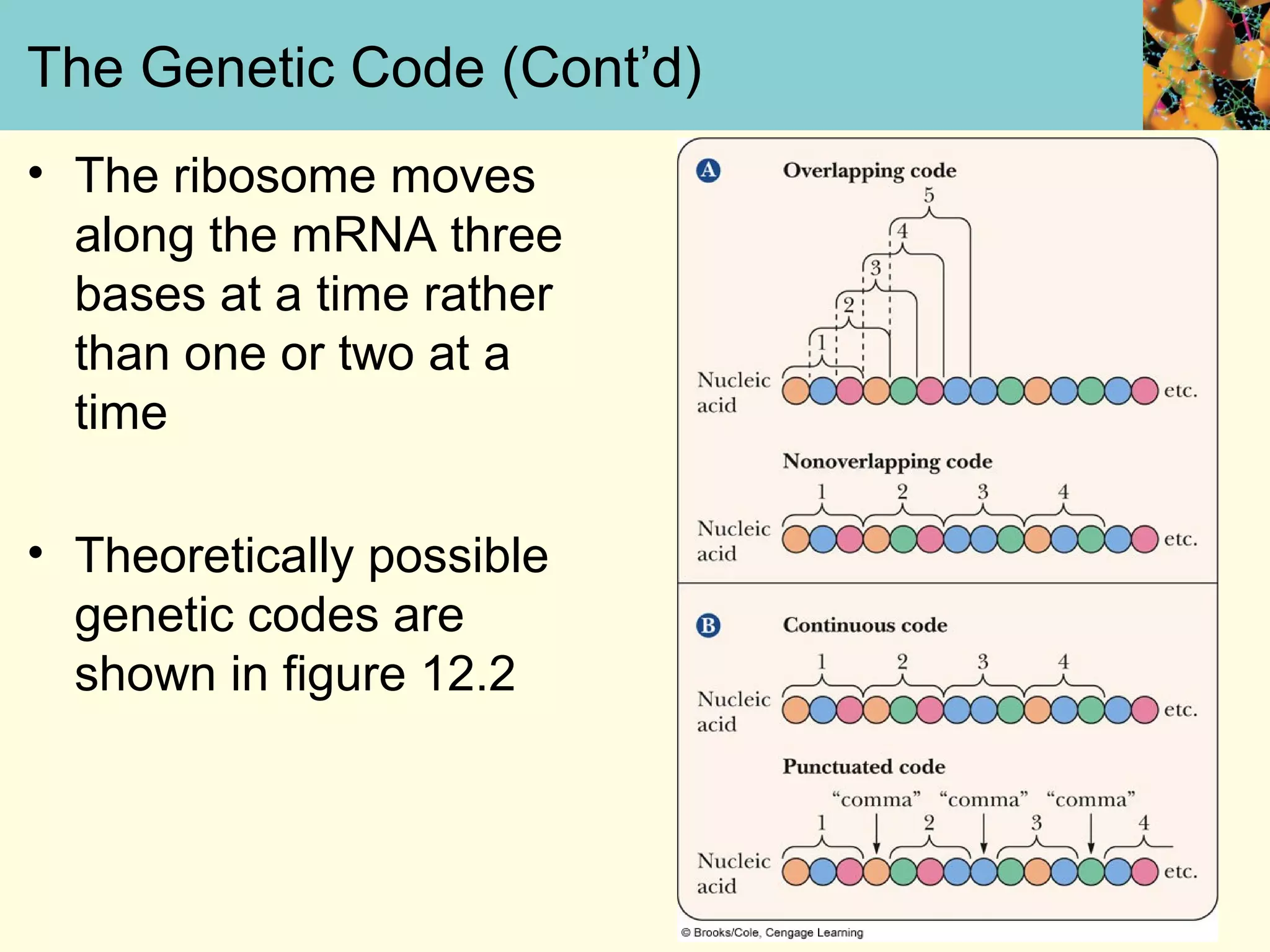
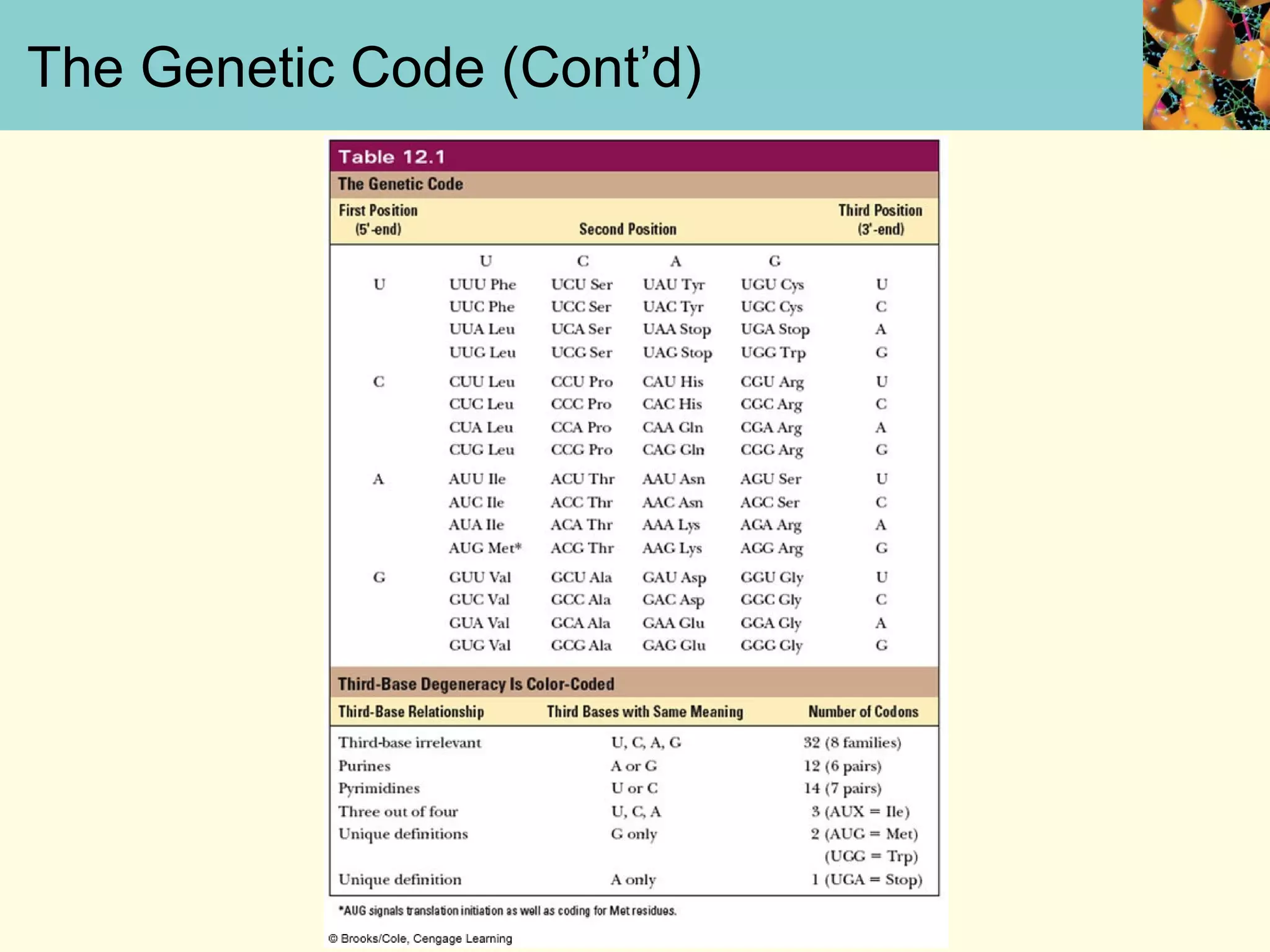
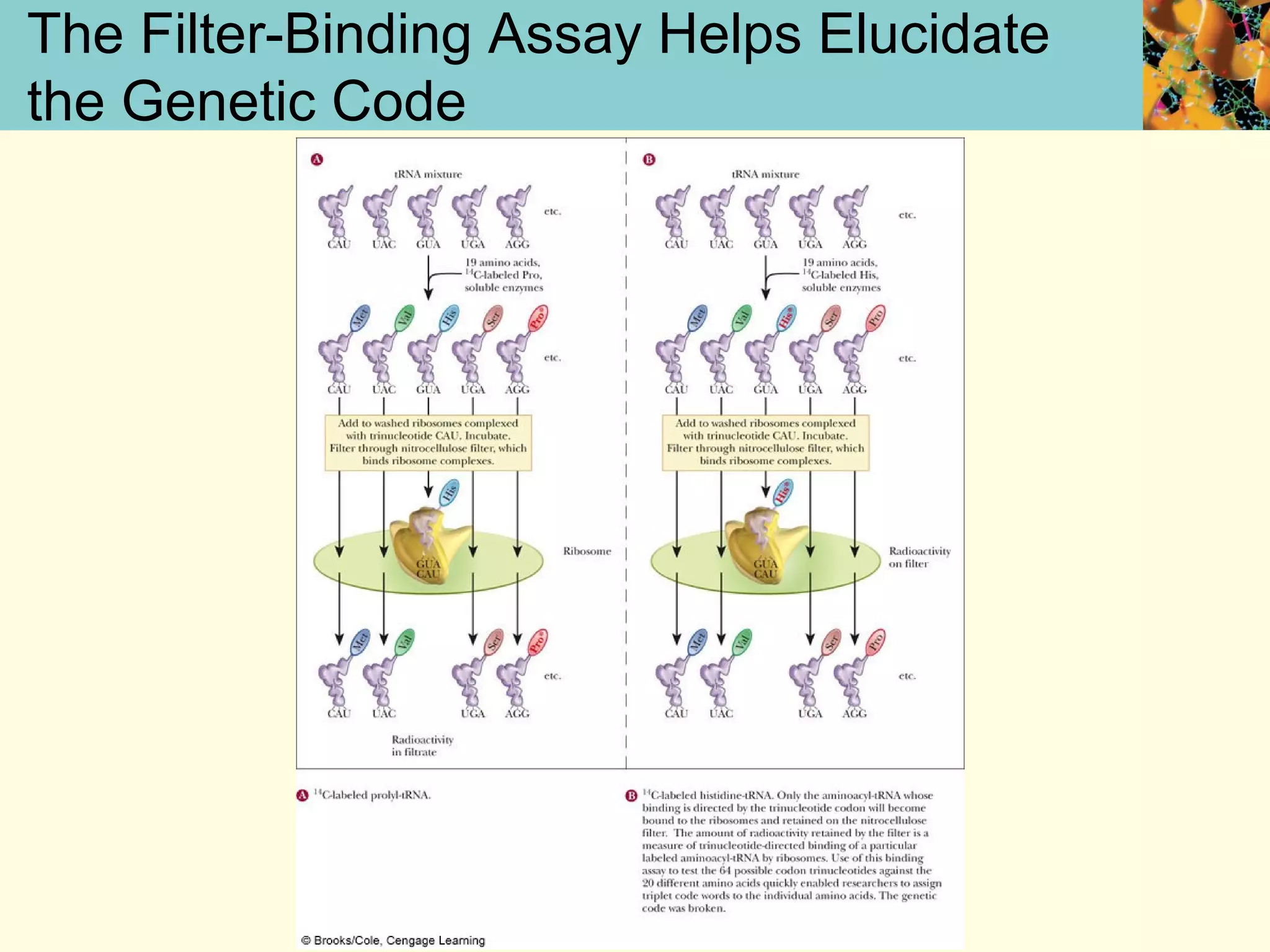
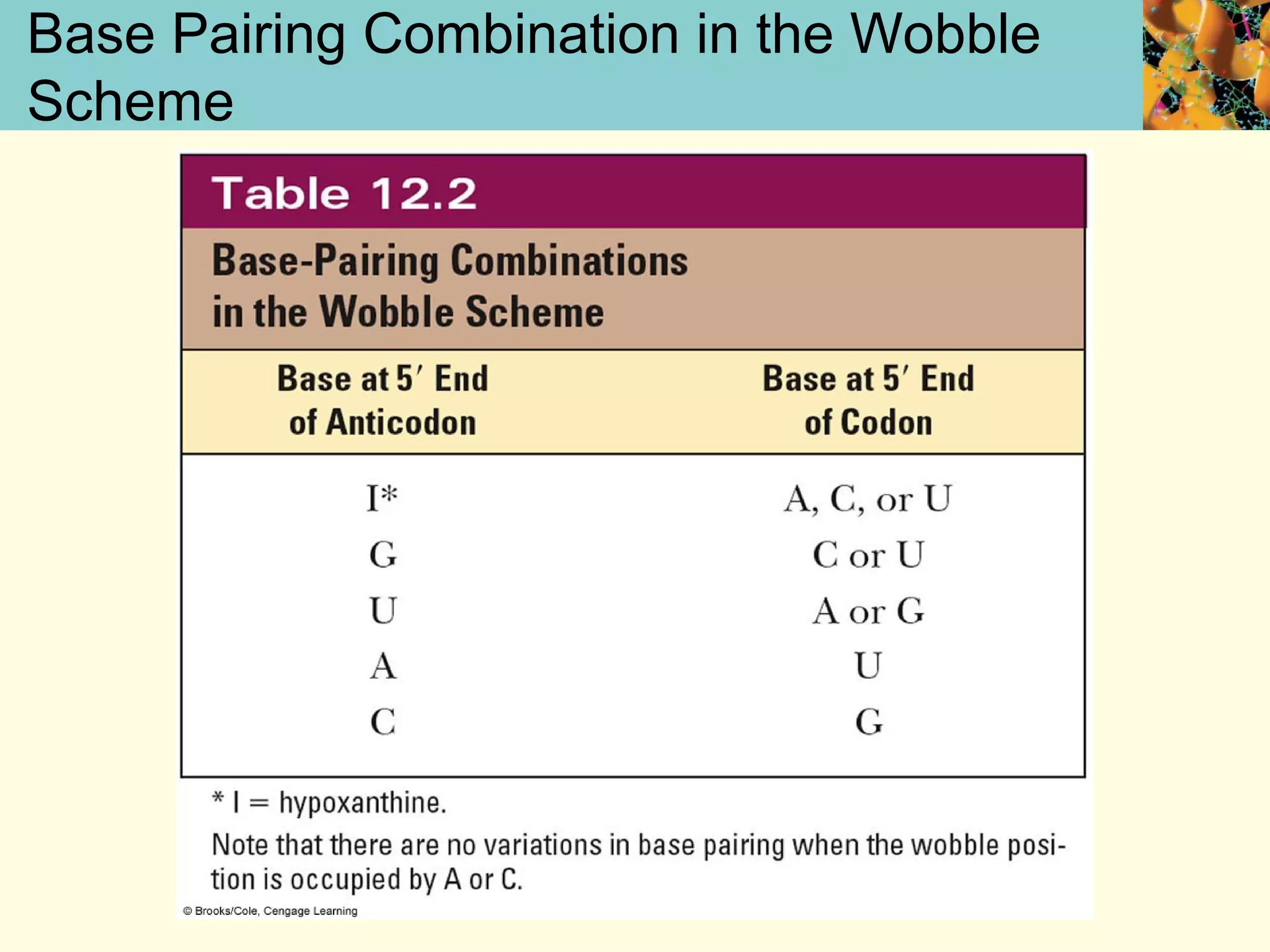
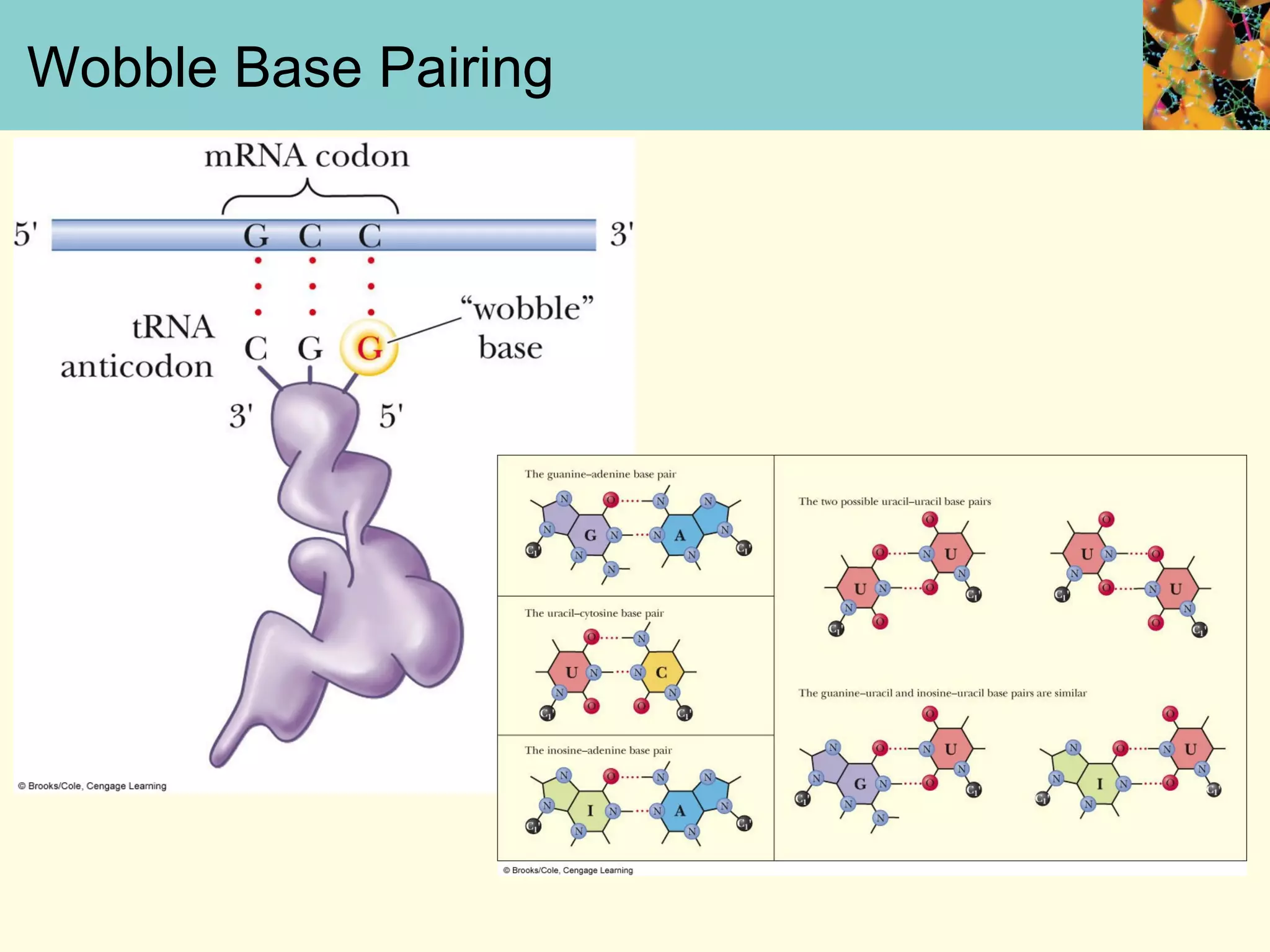
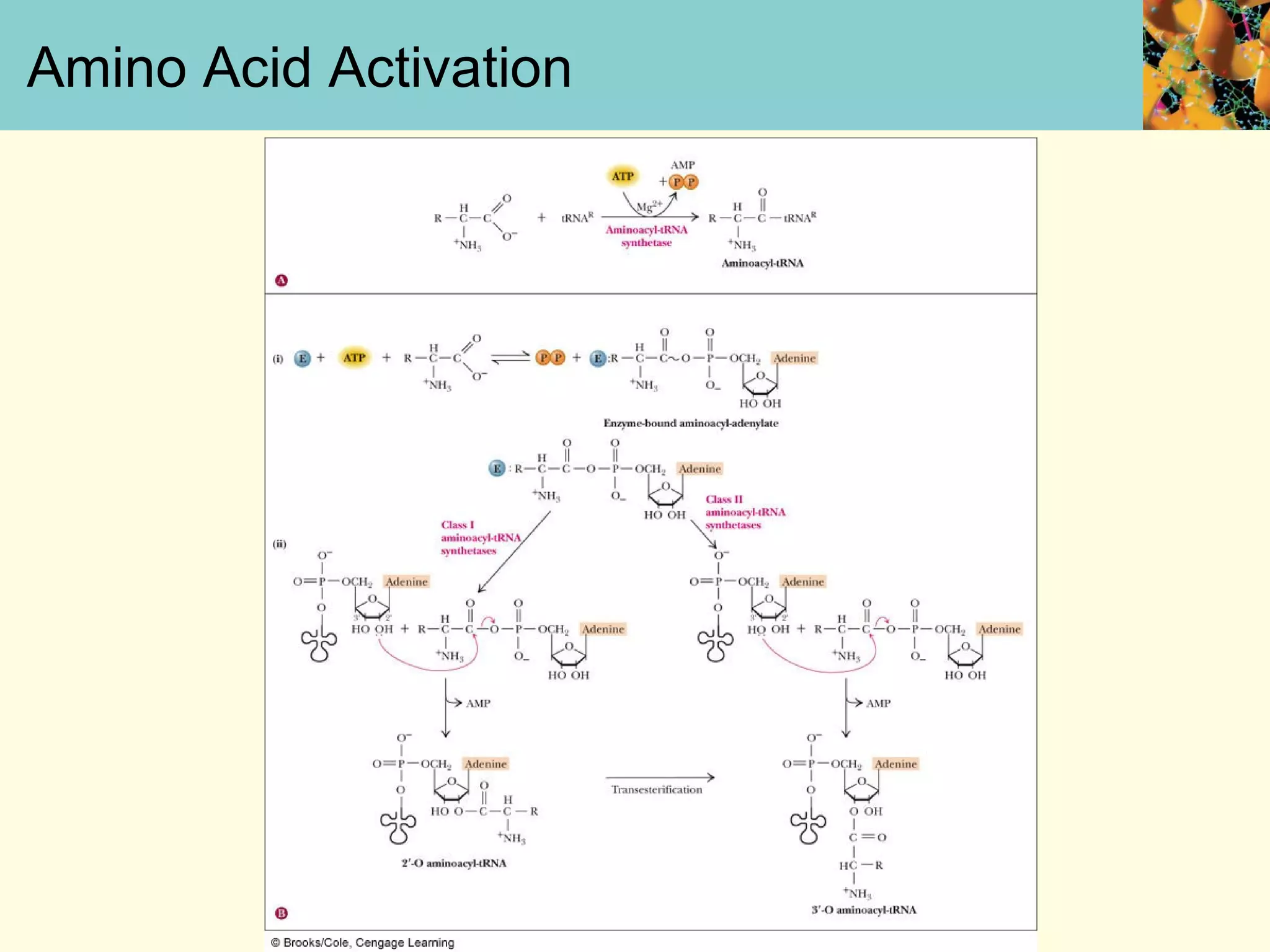
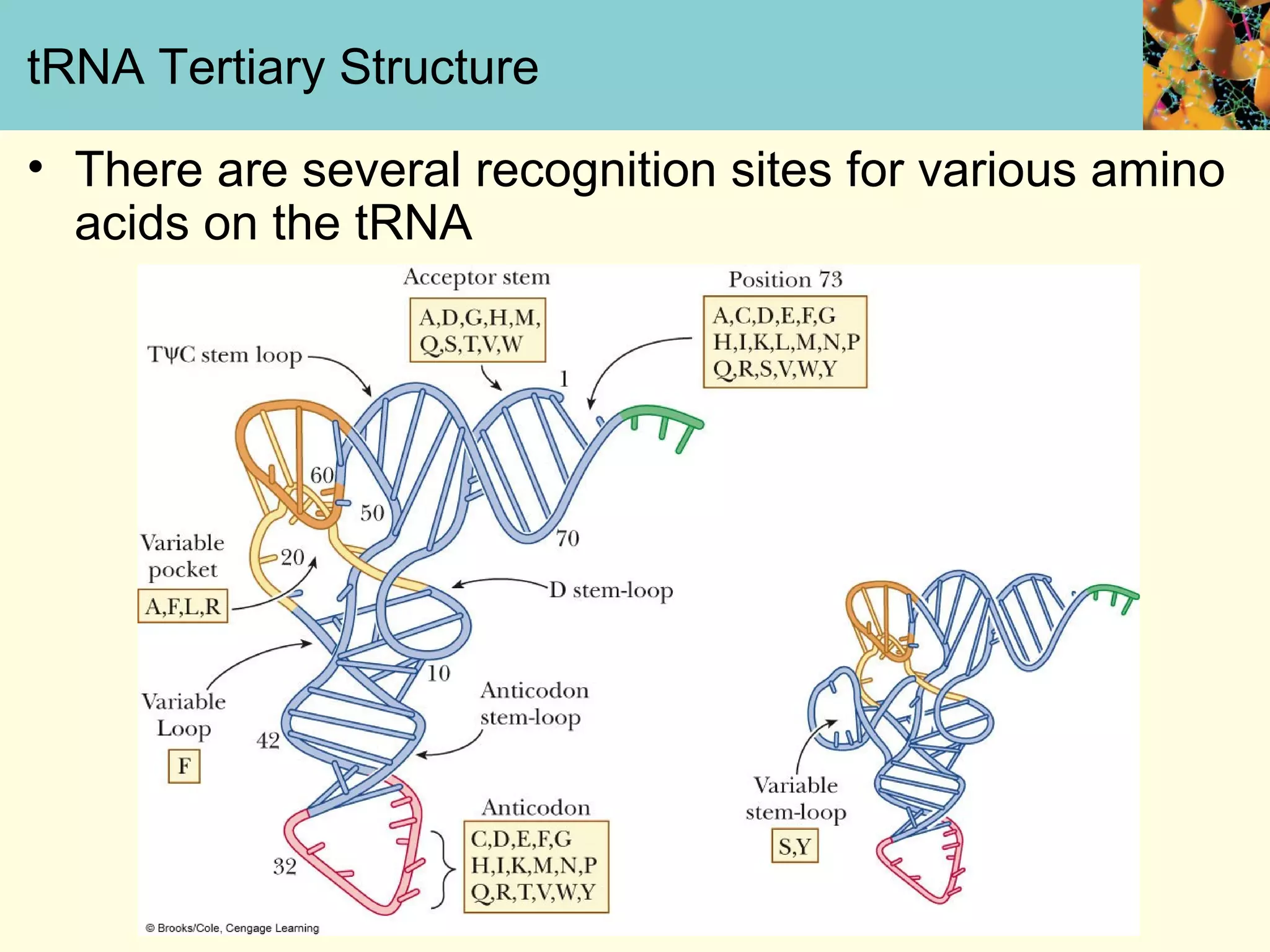
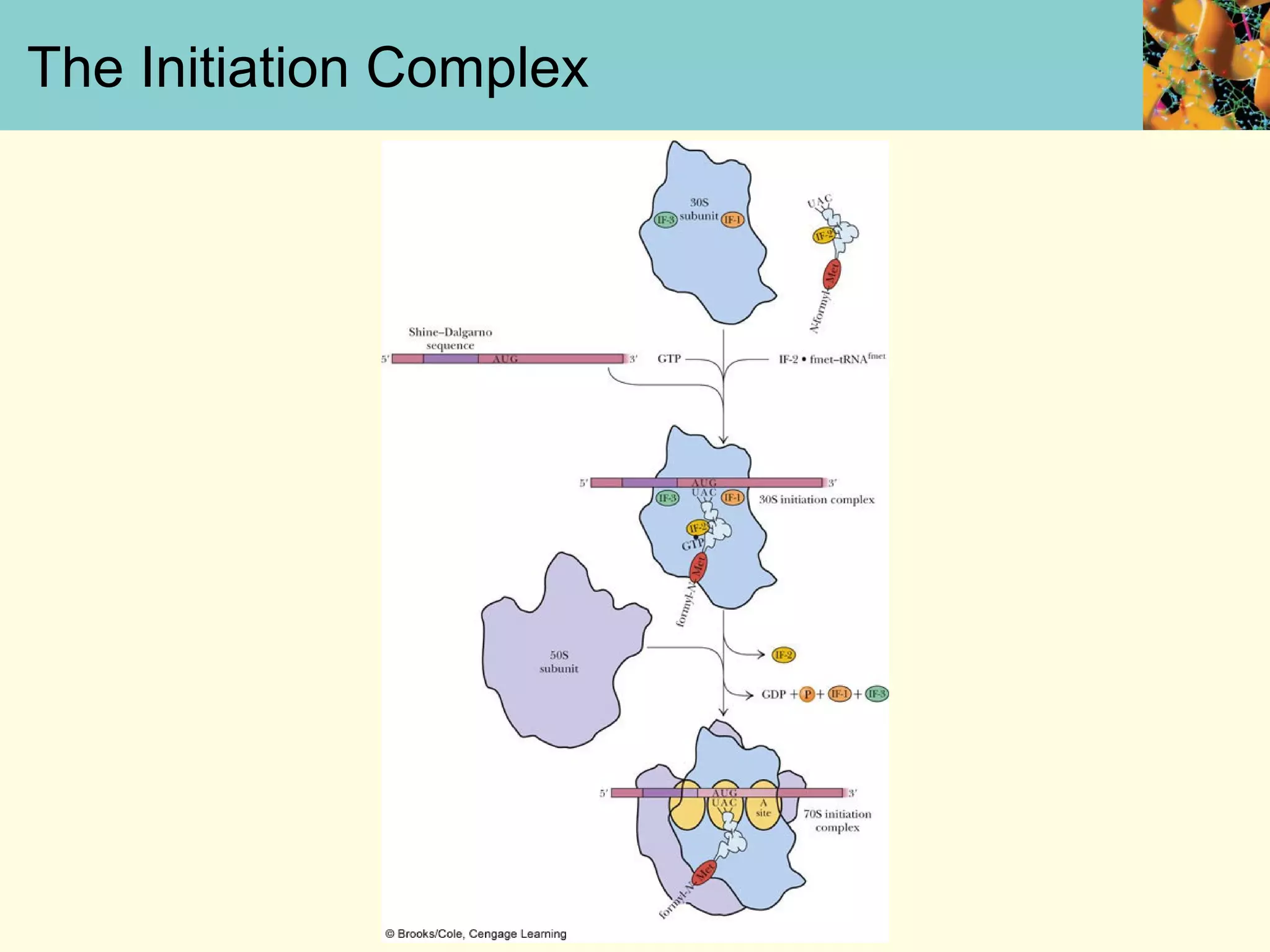
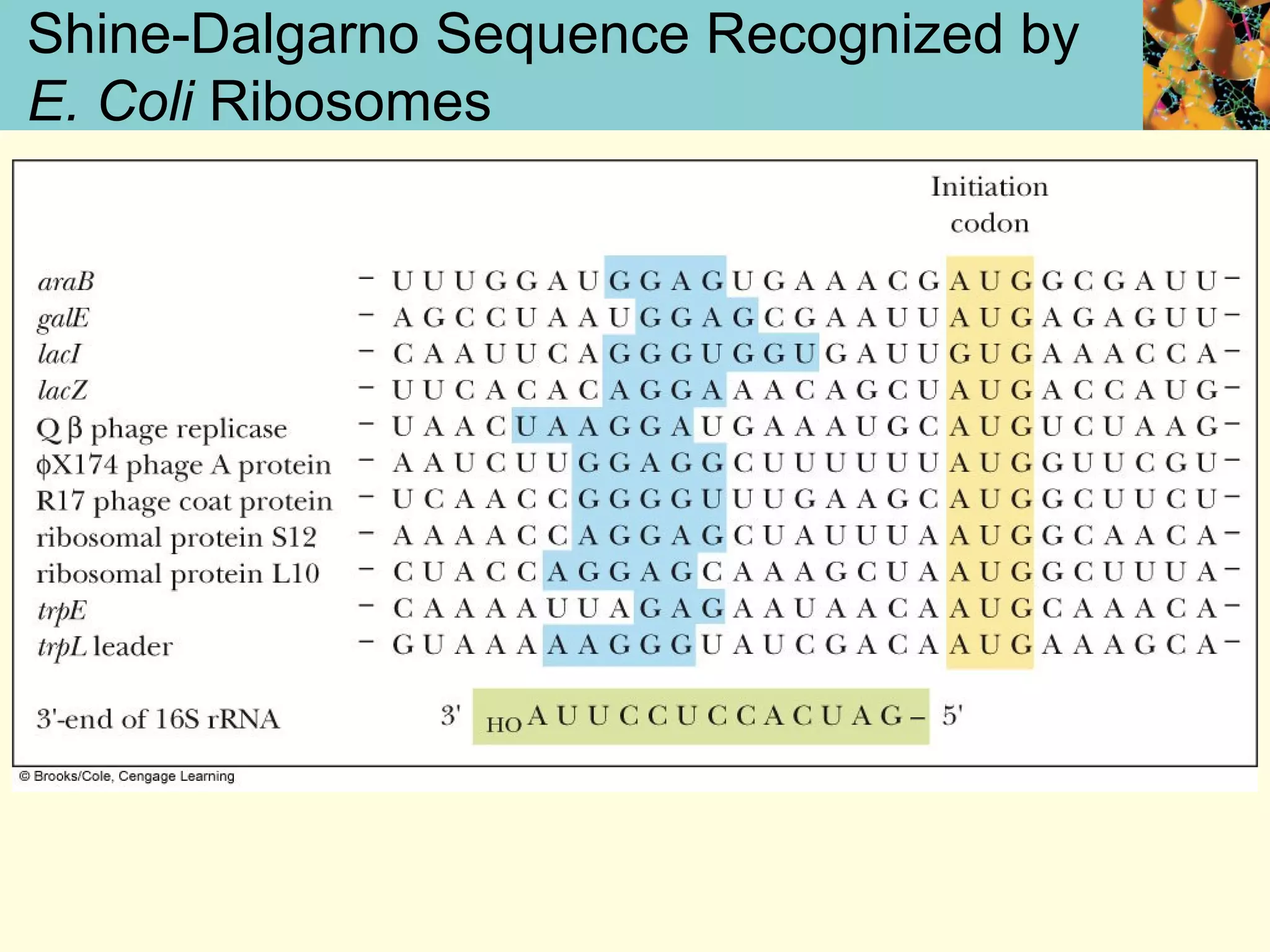
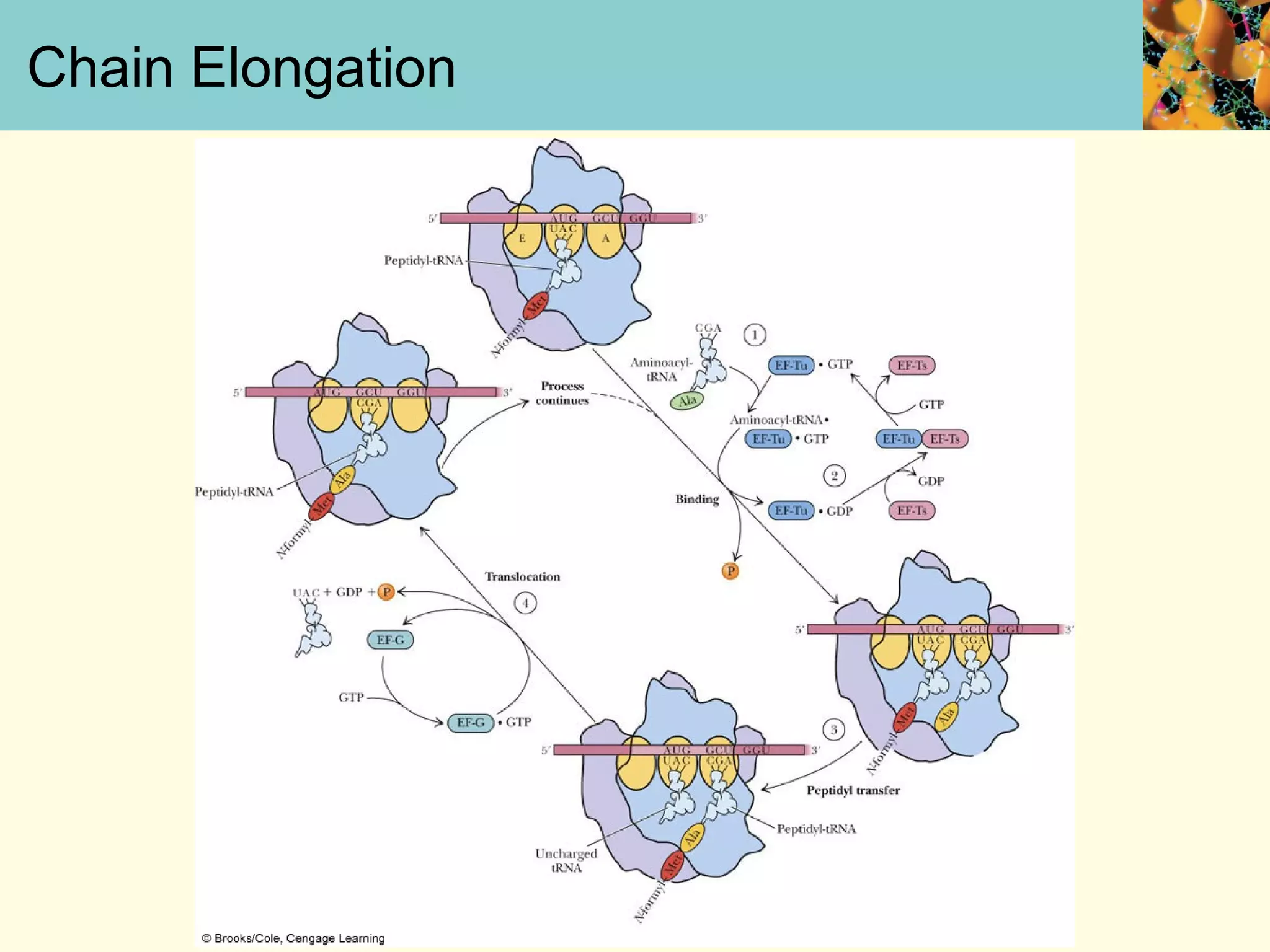
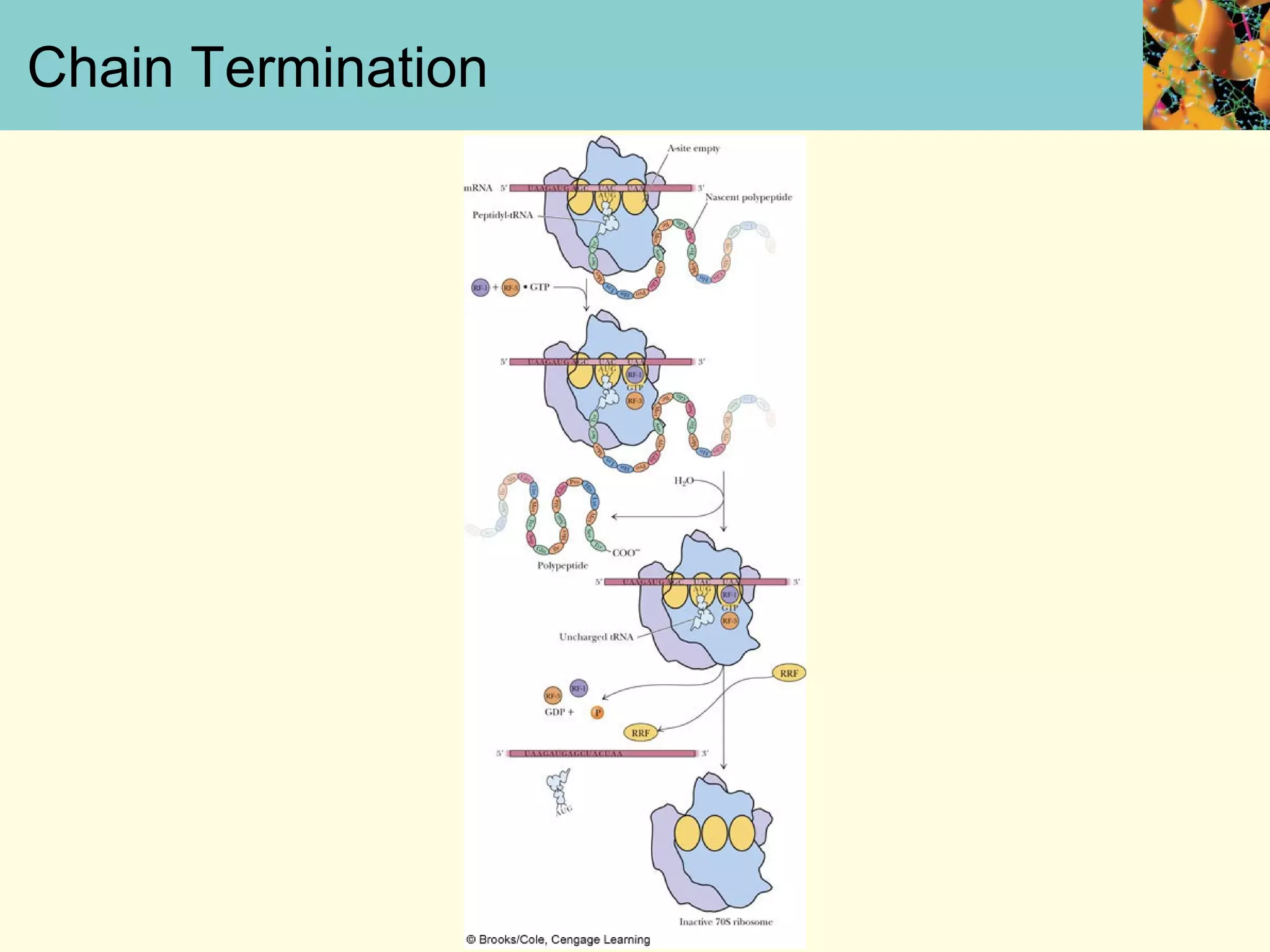
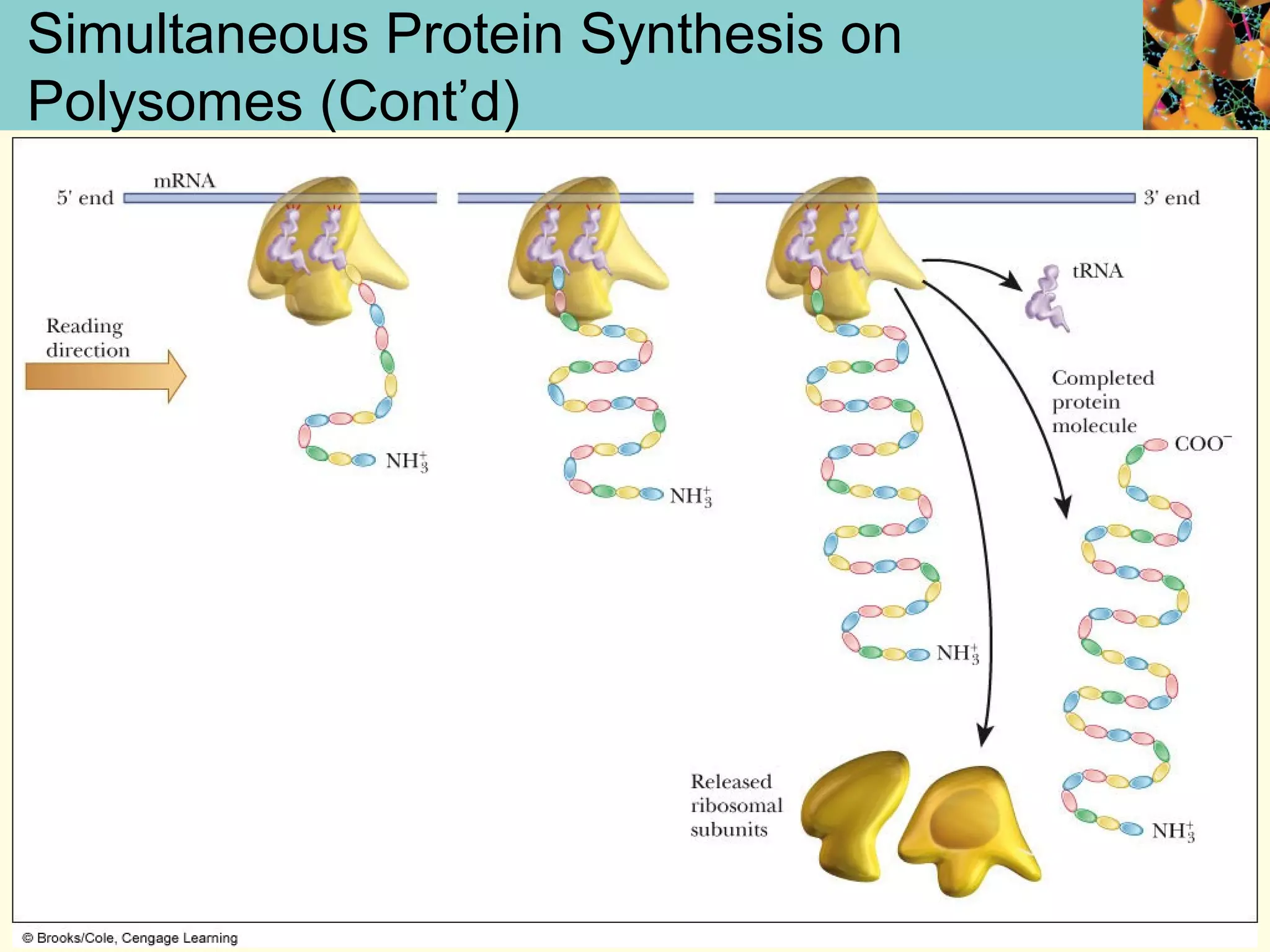
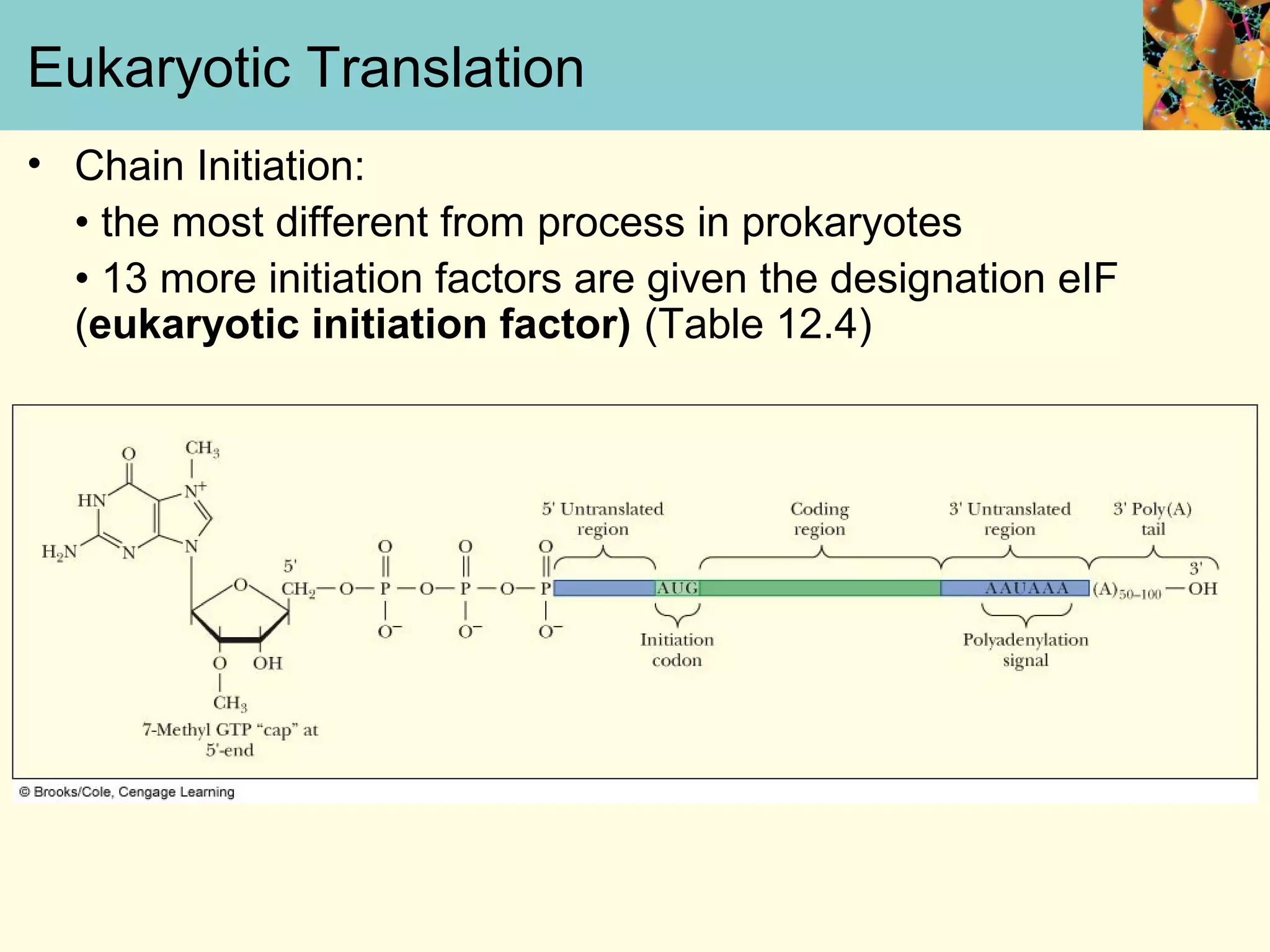
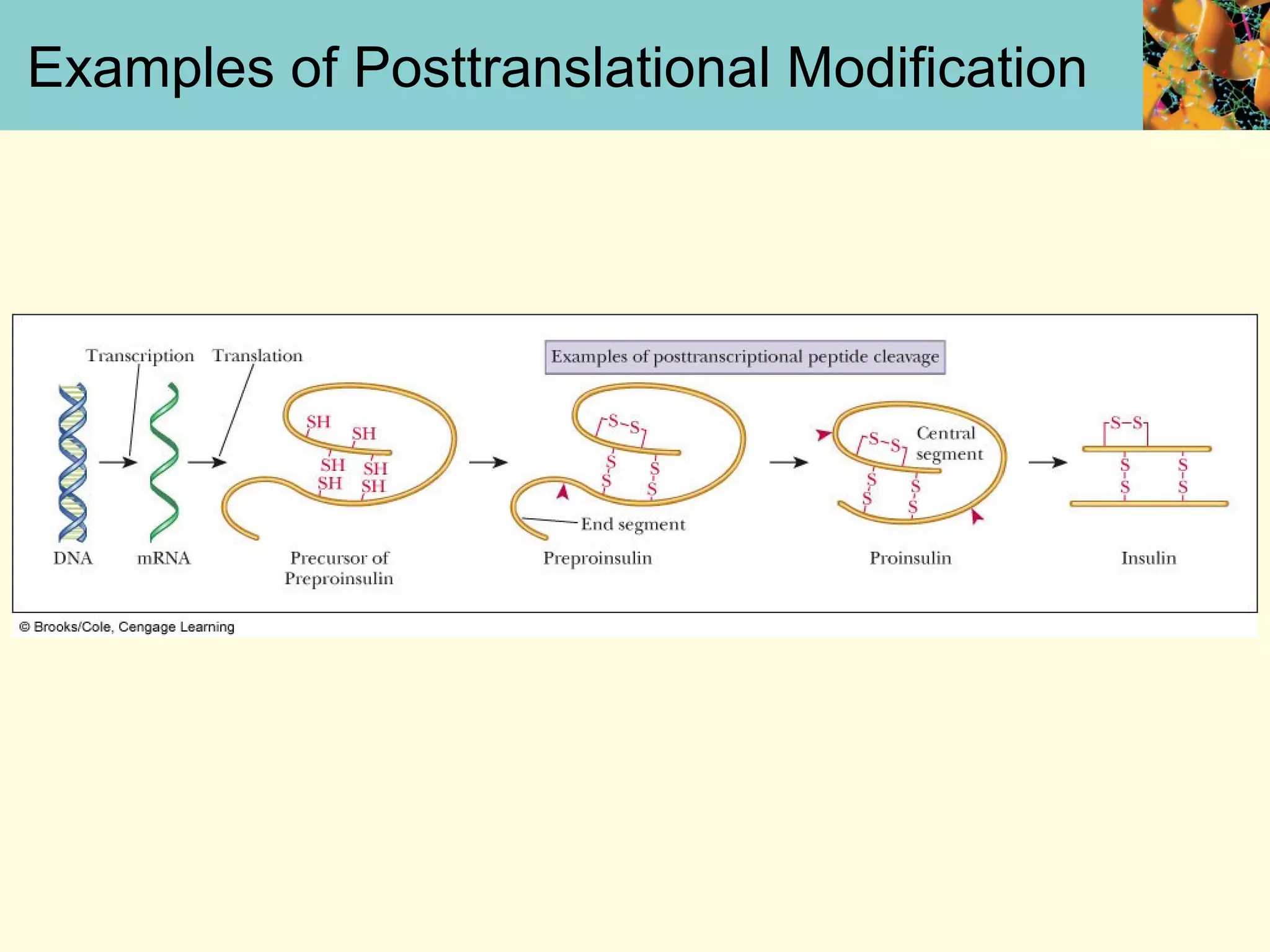
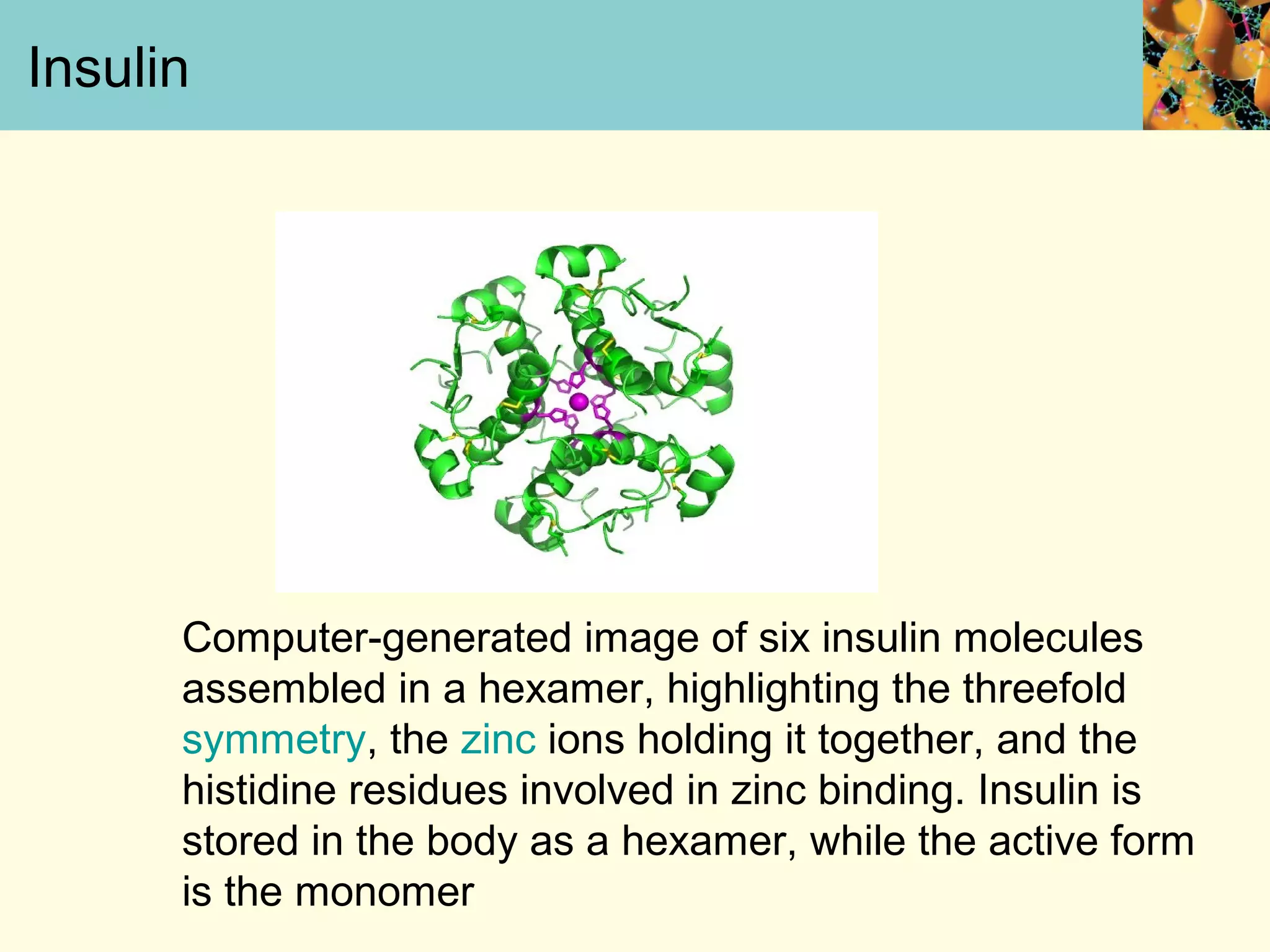
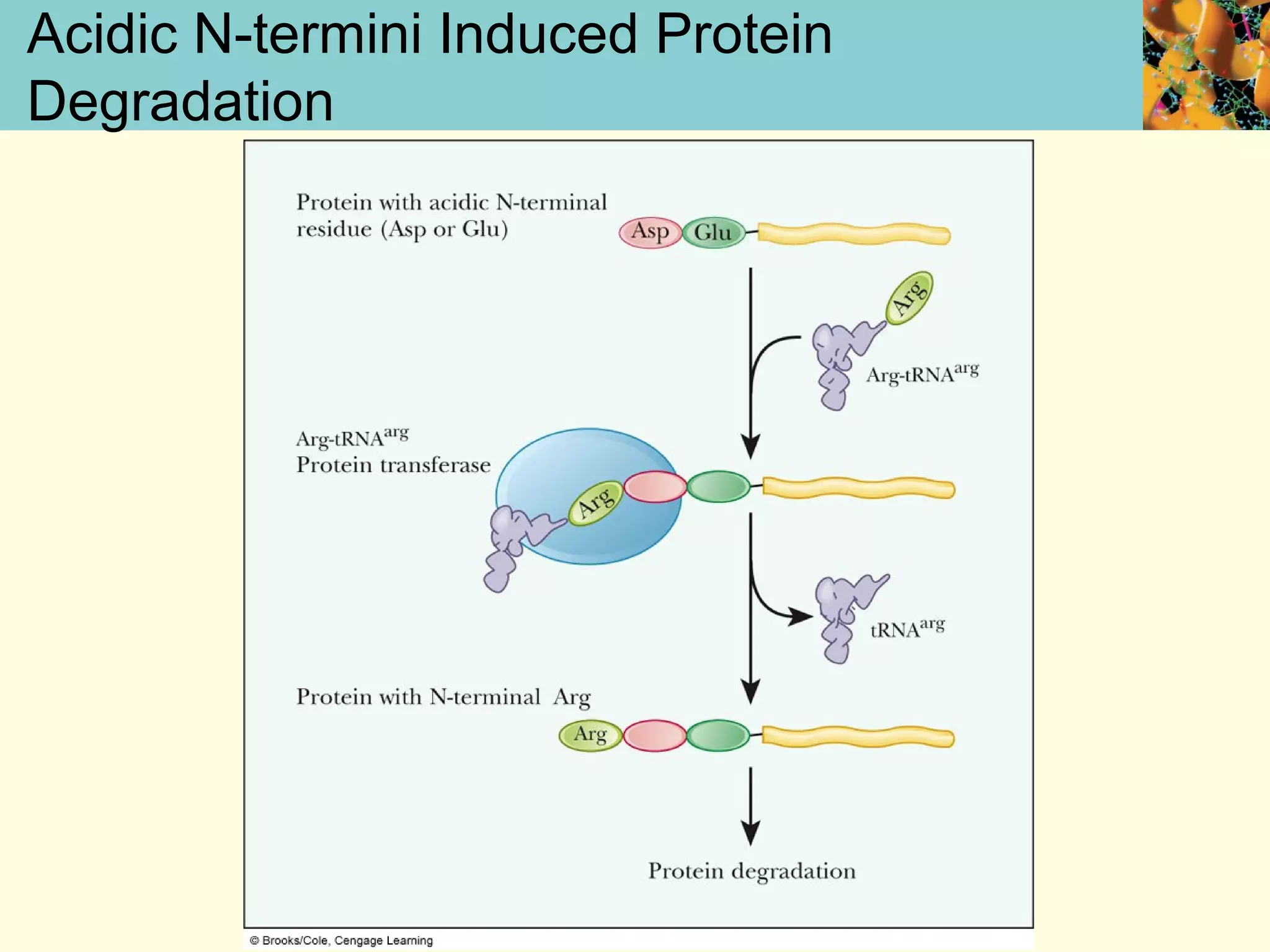
This document summarizes key aspects of protein synthesis, including translation of mRNA into a polypeptide chain. It discusses the genetic code and how triplet codons specify amino acids. The stages of translation - initiation, elongation, and termination - are described. Post-translational modifications and protein degradation are also covered. Protein synthesis requires various ribosomal and transfer RNA components to translate the genetic message into proteins.




































![7 29-10enzymeskinetics-coloso-110715062024-phpapp01[1]](https://cdn.slidesharecdn.com/ss_thumbnails/7-29-10enzymeskinetics-coloso-110715062024-phpapp011-120408174622-phpapp02-thumbnail.jpg?width=640&height=640&fit=bounds)





















































