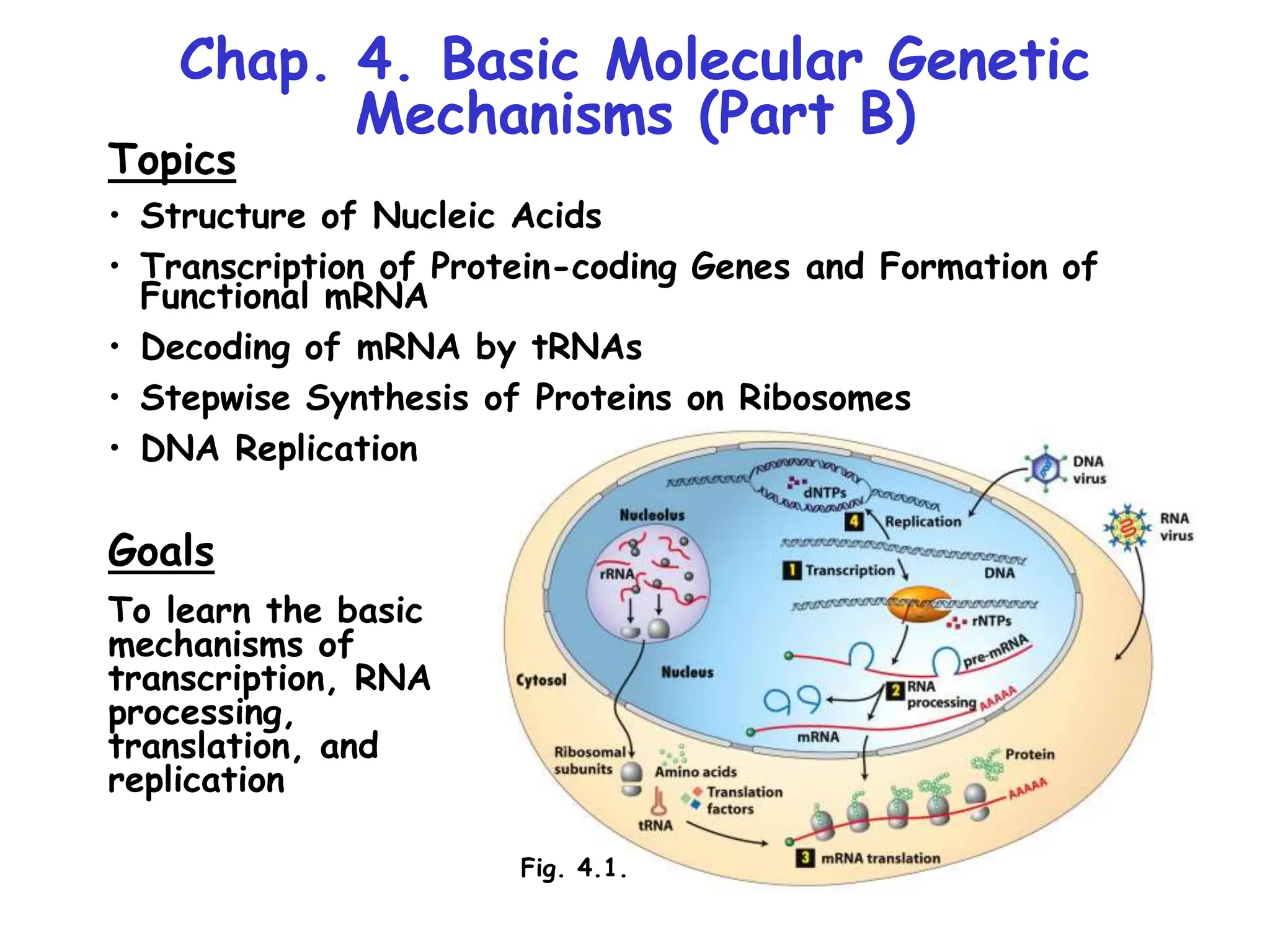
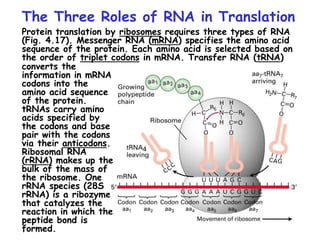
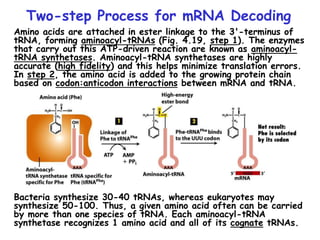
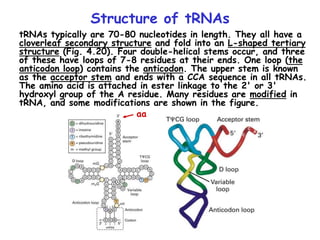
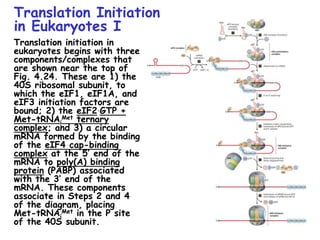
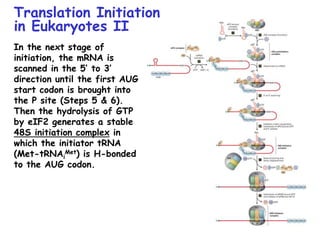
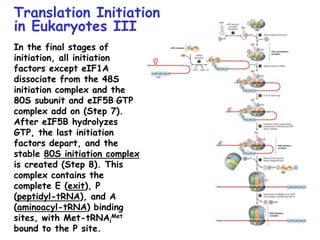
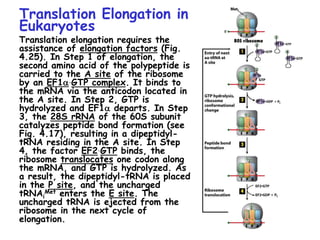
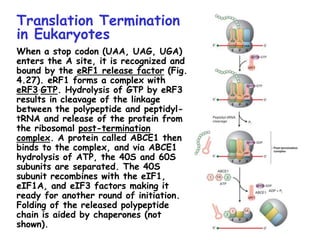
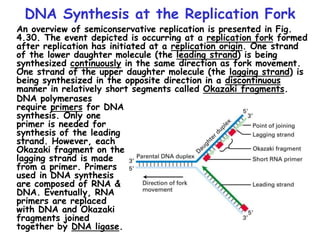
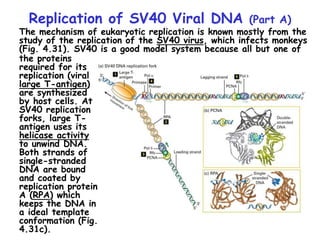
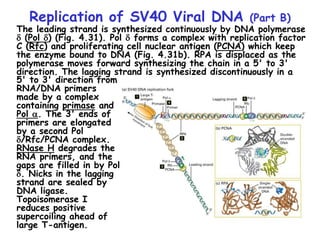
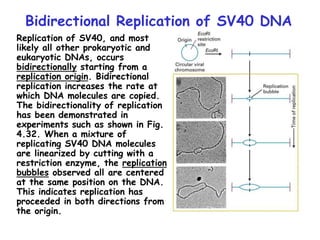
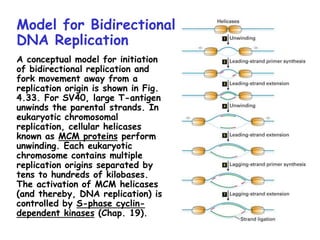
The document summarizes key mechanisms of DNA replication and protein synthesis in cells. It discusses how mRNA is translated into proteins with the help of tRNAs and ribosomes. DNA replication involves unwinding of the DNA double helix at the replication fork and semi-conservative replication of both strands to produce two identical copies of DNA. Bidirectional replication from an origin of replication increases the efficiency of DNA copying.