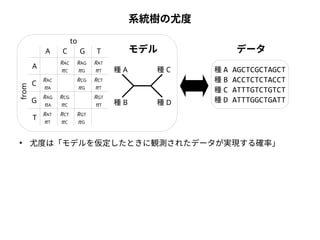
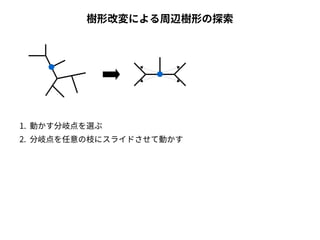
系統樹の尤度
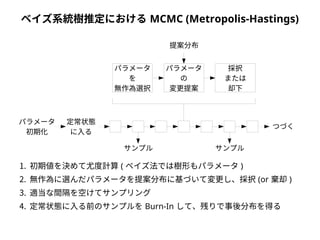
to
from
A
A
C G T
C
G
T
RAC
πC
RAG
πG
RAT
πT
RCG
πC
RCG
πG
RGT
πG
RCT
πT
RGT
πT
RAC
πA
RAG
πA
RAT
πT
RCT
πC
種A
種B
モデルデータ
種C
種D
種A AGCTCGCTAGCT
種B ACCTCTCTACCT
種C ATTTGTCTGTCT
種D ATTTGGCTGATT
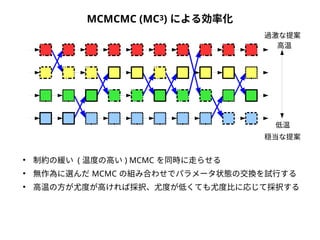
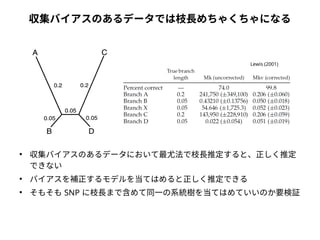
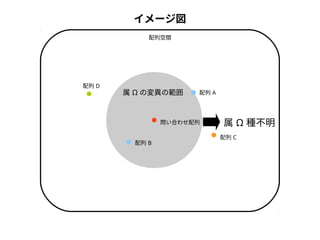
● 尤度は「モデルを仮定したときに観測されたデータが実現する確率」
33.
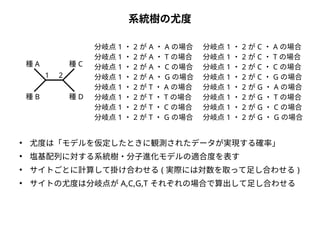
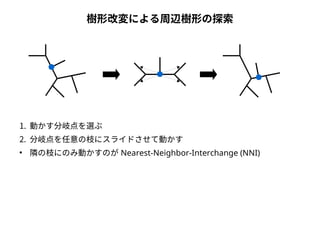
系統樹の尤度
to
from
A
A
C G T
C
G
T
RAC
πC
RAG
πG
RAT
πT
RCG
πC
RCG
πG
RGT
πG
RCT
πT
RGT
πT
RAC
πA
RAG
πA
RAT
πT
RCT
πC
種A
種B
モデルデータ
種C
種D
種A AGCTCGCTAGCT
種B ACCTCTCTACCT
種C ATTTGTCTGTCT
種D ATTTGGCTGATT
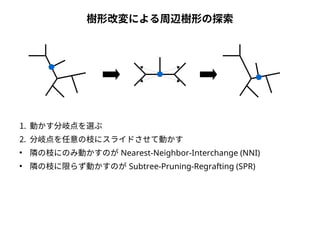
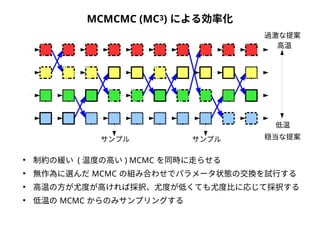
● 尤度は「モデルを仮定したときに観測されたデータが実現する確率」
● 塩基配列に対する系統樹・分子進化モデルの適合度を表す
34.
系統樹の尤度
種A AG C T C G C T A G C T
種B A C C T C T C T A C C T
種C A T T T G T C T G T C T
種D A T T T G G C T G A T T
lnL -5 -3 -9 -8 -5 -7 -5 -5 -4 -6 -4 -5
● 尤度は「モデルを仮定したときに観測されたデータが実現する確率」
● 塩基配列に対する系統樹・分子進化モデルの適合度を表す
● サイトごとに計算して掛け合わせる( 実際には対数を取って足し合わせる)
系統樹の尤度
to
from
A
A
C G T
C
G
T
RAC
πC
RAG
πG
RAT
πT
RCG
πC
RCG
πG
RGT
πG
RCT
πT
RGT
πT
RAC
πA
RAG
πA
RAT
πT
RCT
πC
種A
種B
モデルデータ
種C
種D
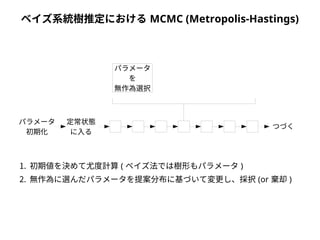
● 尤度は「モデルを仮定したときに観測されたデータが実現する確率」
● 塩基配列に対する系統樹・分子進化モデルの適合度を表す
● サイトごとに計算して掛け合わせる( 実際には対数を取って足し合わせる)
● サイトの尤度は分岐点がA,C,G,T それぞれの場合で算出して足し合わせる
● RXY・πX・枝長は数値計算により最適化
種A AGCTCGCTAGCT
種B ACCTCTCTACCT
種C ATTTGTCTGTCT
種D ATTTGGCTGATT
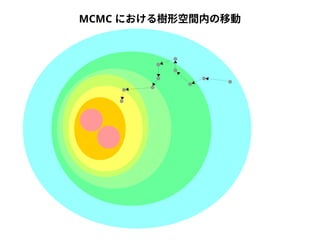
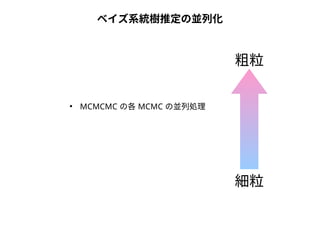
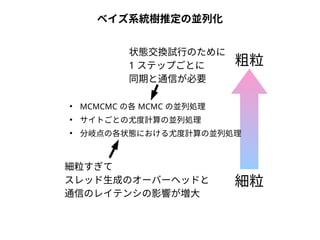
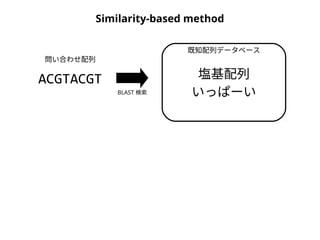
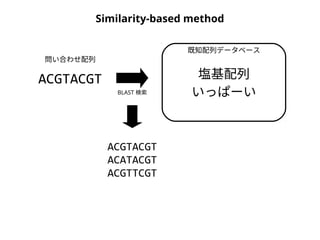
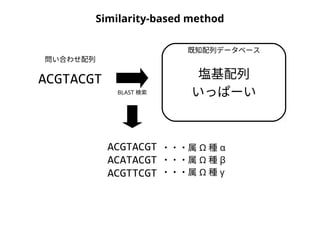
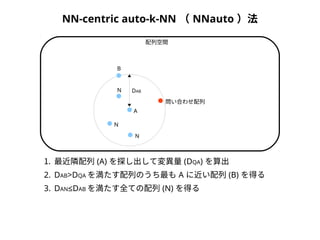
NN-centric auto-k-NN (NNauto )法
DQA
A
B
配列空間
問い合わせ配列
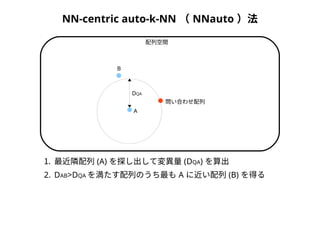
1. 最近隣配列(A) を探し出して変異量(DQA) を算出
2. DAB>DQAを満たす配列のうち最もA に近い配列(B) を得る
186.
NN-centric auto-k-NN (NNauto )法
DAB
A
B
N
N
N
配列空間
問い合わせ配列
1. 最近隣配列(A) を探し出して変異量(DQA) を算出
2. DAB>DQAを満たす配列のうち最もA に近い配列(B) を得る
3. DAN≤DABを満たす全ての配列(N) を得る
187.
NN-centric auto-k-NN (NNauto )法
DAB
A
B
N
N
N
配列空間
問い合わせ配列
1. 最近隣配列(A) を探し出して変異量(DQA) を算出
2. DAB>DQAを満たす配列のうち最もA に近い配列(B) を得る
3. DAN≤DABを満たす全ての配列(N) を得る
4. A, B, N の全配列で共通する分類群を採用
188.
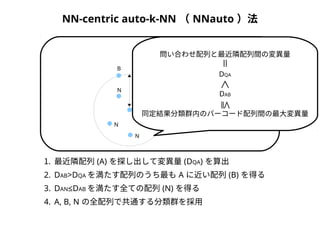
NN-centric auto-k-NN (NNauto )法
DAB
A
B
N
N
N
配列空間
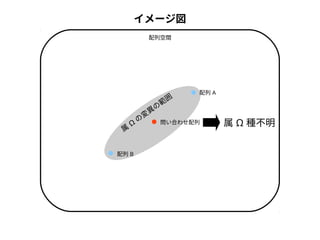
問い合わせ配列と最近隣配列間の変異量
問い合わせ配列
< ≦ =
DQA
DAB
同定結果分類群内のバーコード配列間の最大変異量
1. 最近隣配列(A) を探し出して変異量(DQA) を算出
2. DAB>DQAを満たす配列のうち最もA に近い配列(B) を得る
3. DAN≤DABを満たす全ての配列(N) を得る
4. A, B, N の全配列で共通する分類群を採用
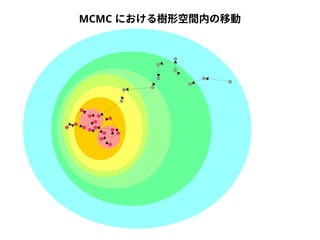
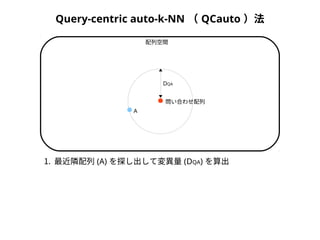
Query-centric auto-k-NN (QCauto )法
DQA
A
B
配列空間
問い合わせ配列
1. 最近隣配列(A) を探し出して変異量(DQA) を算出
2. DAB>DQAを満たす配列のうち最もA に近い配列(B) を得る
192.
Query-centric auto-k-NN (QCauto )法
A
B
N
N
配列空間
DQB
問い合わせ配列
N
1. 最近隣配列(A) を探し出して変異量(DQA) を算出
2. DAB>DQAを満たす配列のうち最もA に近い配列(B) を得る
3. DQN≤DQBを満たすすべての配列(N) を得る
193.
Query-centric auto-k-NN (QCauto )法
A
B
N
N
配列空間
DQB
問い合わせ配列
N
1. 最近隣配列(A) を探し出して変異量(DQA) を算出
2. DAB>DQAを満たす配列のうち最もA に近い配列(B) を得る
3. DQN≤DQBを満たすすべての配列(N) を得る
4. A, B, N の全配列で共通する分類群を採用
194.
Query-centric auto-k-NN (QCauto )法
A
B
N
N
配列空間
問い合わせ配列と最近隣配列間の変異量
DQB
問い合わせ配列
< ≦ =
同定結果分類群内のバーコード配列間の最大変異量
N
DQA
DQB
1. 最近隣配列(A) を探し出して変異量(DQA) を算出
2. DAB>DQAを満たす配列のうち最もA に近い配列(B) を得る
3. DQN≤DQBを満たすすべての配列(N) を得る
4. A, B, N の全配列で共通する分類群を採用
























































































































































































































![[PRML] パターン認識と機械学習(第1章:序論)](https://cdn.slidesharecdn.com/ss_thumbnails/prmlchapter1-170903070406-thumbnail.jpg?width=640&height=640&fit=bounds)



















