
Recommended
Recommended
More Related Content
What's hot
What's hot (20)
More from jlquinonesrivera
More from jlquinonesrivera (13)
Recently uploaded
Recently uploaded (20)
DNA- replicación - transcripción
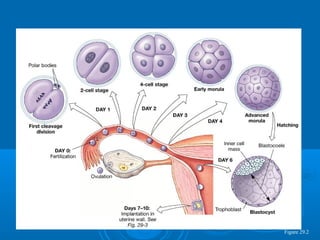
- 1. Figure 29.2
- 3. Griffith’s Transformation ExperimentGriffith’s Transformation Experiment 19281928
- 6. Animals Human Homo sapiens 46 Chimpanzee Pan troglodytes 48 Gorilla 48 Orangutan Pongo x 48 Rhesus Monkey 42 Mouse Mus musculus 40 Cat Felis catus 38 Pig 38 Wolf Canis lupus 78 Dog Canis lupus familiaris 78 Cow Bos primigenius 60 Goat 60 Plants Oats Avena sativa 42 Mango Mangifera indica 40 Rice Oryza sativa 24 Bean Phaseolus sp. 22 Wheat Triticum aestivum 42
- 11. James Watson, Francis Crick, and Maurice Wilkins received a Nobel Prize for the double-helix model of DNA in 1962, four years after Rosalind Franklin's death at age 37 from ovarian cancer.
- 17. A-DNAconformation B-DNA conformation Z-DNA conformation
- 20. Fig. 10-2, p. 213
- 24. Fig. 10-4, p. 214
- 26. Fig. 10-3, p. 213 DNA Scaffolding proteins 2 μm
- 30. Fig. 12-7a, p. 268 (a) Hypothesis 1: Semiconservative replication Parental DNA First generation Second generation
- 32. Replication in Bacteria andReplication in Bacteria and EukaryotesEukaryotes
- 44. DNA Síntesis
- 46. E. coli DNA polymerase III
- 47. Fig. 12-15a, p. 276 DNA replication RNA primer RNA primer Removal of primer 3’ 3’ 3’ 3’ 3’ 3’ 3’ 3’ 3’ 3’ 5’ 5’ 5’ 5’ 5’ 5’ 5’ 5’ 5’ 5’ + +
- 49. Miles depurinaciones diarias 100 deaminaciones diarias
- 55. Fig. 13-4, p. 283 Nontemplate strand Transcription DNA Template strand mRNA (complementary copy of template DNA strand) Codon 1 Codon 2 Codon 3 Codon 4 Codon 5 Codon 6 Polypeptide Met Thr Cys Glu Cys Phe Translation ‘ ‘ ‘ ‘ ‘ ‘
- 101. enzima de restricción Hpa II
- 105. Transcripción
Editor's Notes
- Figure 19.15 B-DNA. (a) Ball-and-stick model. The base pairs are nearly perpendicular to the sugar– phosphate backbones. (b) Space-filling model. Color key: carbon, gray; nitrogen, blue; oxygen, red; phosphorus, purple. [Nucleic Acids Database BD0001].
- Figure 19.7 Chemical structures of nucleosides. The carbon atoms of the sugars are numbered with primes to distinguish them from the atoms of the bases. (a) Ribonucleosides. The sugar in ribonucleosides is ribose, which contains a hydroxyl group at C-2', as shown here. The -N-glycosidic bond of adenosine is shown in red. (b) Deoxyribonucleosides. In deoxyribonucleosides, there is an additional hydrogen atom at C-2' instead of a hydroxyl group.
- Figure 19.9 Chemical structures of the deoxyribonucleoside- 5'-monophosphates.
- Figure 19.11 Chemical structure of the tetranucleotide pdApdGpdTpdC. The nucleotide residues are joined by 3'—5’ phosphodiester linkages. The nucleotide with a free 5'-phosphoryl group is called the 5' end, and the nucleotide with a free 3'-hydroxyl group is called the 3’ end.
- Figure 19.12 Chemical structure of double-stranded DNA. The two strands run in opposite directions. Adenine in one strand pairs with thymine in the opposite strand, and guanine pairs with cytosine.
- Figure 19.18 The A-DNAconformation (left) is favored when DNA is dehydrated [NDB AD0001]. B-DNA (center) is the conformation normally found inside cells [NDB BD0001]. The Z-DNA conformation (right) is favored in certain G/C-rich sequences [NDB ZDJ050].
- Figure 10.2: Nucleosomes.
- Figure 19.23 Diagram of nucleosome structure. (a) Histone octamer. (b) Nucleosomes. Each nucleosome is composed of a core particle plus histone H1 and linker DNA. The nucleosome core particle is composed of a histone octamer and about 146 bp of DNA. Linker DNA consists of about 54 bp. Histone H1 binds to the core particle and to linker DNA.
- Figure 19.23 Diagram of nucleosome structure. (a) Histone octamer. (b) Nucleosomes. Each nucleosome is composed of a core particle plus histone H1 and linker DNA. The nucleosome core particle is composed of a histone octamer and about 146 bp of DNA. Linker DNA consists of about 54 bp. Histone H1 binds to the core particle and to linker DNA.
- Figure 19.25 A model of the 30 nm chromatin fiber. In this model the 30 nm fiber is shown as a solenoid, or helix, formed by individual nucleosomes. The nucleosomes associate through contacts between adjacent histone H1 molecules.
- Figure 10.4: Organization of a eukaryotic chromosome. This diagram shows how DNA is packaged into highly condensed chromosomes. First, DNA is wrapped around histone proteins to form nucleosomes. Then, the nucleosomes are compacted into chromatin fibers, which are coiled into looped domains. The looped domains are compacted, ultimately forming chromosomes.
- Figure 10.3: TEM of a mouse chromosome depleted of histones. Notice how densely packed the DNA strands are, even though they have been released from the histone proteins that organize them into tightly coiled structures.
- Figure 12.7: Alternative models of DNA replication. The hypothesized arrangement of old (light blue) and newly synthesized (dark blue) DNA strands after one and two generations, according to (a) the semiconservative model, (b) the conservative model, and (c) the dispersive model.
- Figure 20.2 Bidirectional DNA replication in Escherichia coli. Semiconservative DNA replication begins at a unique origin and proceeds in both directions. The synthesis of new strands of DNA (light gray) occurs at the two replication forks, where the replisomes are located. When the replication forks meet at the termination site, the two double-stranded DNA molecules separate. Note that each daughter molecule consists of one parental strand and one newly synthesized strand.
- Figure 20.5 Diagram of the subunit composition of E. coli DNA polymerase III. The holoenzyme consists of two core complexes (containing , , and ), paired copies of and and a single complex (3,,’, with two copies each of and ). The structure is thus an asymmetric dimer. Other models of the holoenzyme structure have been proposed. [Adapted from O’Donnell, M. (1992). Accessory protein function in the DNA polymerase III holoenzyme from E. coli. BioEssays 14:105–111.]
- Figure 12.15: Replication at chromosome ends.
- Figure 20.21 General excision-repair pathway.
- Figure 20.20 Repair of thymine dimers by DNA photolyase.
- Figure 20.24 Repair of damage resulting from the deamination of cytosine.
- Figure 21.1 Biological information flow. The normal flow of biological information is from DNA to RNA to protein.
- Figure 13.4: An overview of transcription and translation. In transcription, messenger RNA is synthesized as a complementary copy of one of the DNA strands, the template strand. Messenger RNA carries genetic information in the form of sets of three bases, or codons, each of which specifies one amino acid. Codons are translated consecutively, thus specifying the linear sequence of amino acids in the polypeptide chain. Translation requires tRNA and ribosomes (not shown). The figure depicts transcription and translation in bacteria. In eukaryotes, transcription takes place in the nucleus and translation occurs in the cytosol.
- Figure 19.7 Chemical structures of nucleosides. The carbon atoms of the sugars are numbered with primes to distinguish them from the atoms of the bases. (a) Ribonucleosides. The sugar in ribonucleosides is ribose, which contains a hydroxyl group at C-2', as shown here. The -N-glycosidic bond of adenosine is shown in red. (b) Deoxyribonucleosides. In deoxyribonucleosides, there is an additional hydrogen atom at C-2' instead of a hydroxyl group.
- Figure 21.5 Orientation of a gene. The sequence of a hypothetical gene and the RNA transcribed from it are shown. By convention, the gene is said to be transcribed from the 5' end to the 3' end, but the template strand of DNA is copied from the 3' end to the 5' end. Growth of the ribonucleotide chain proceeds 5'-->3'.
- Figure 21.4 Transcription of E. coli ribosomal RNA genes. The genes are being transcribed from left to right. The nascent rRNA product associates with proteins and is processed by nucleolytic cleavage before transcription is complete.
- Figure 21.3 Reaction catalyzed by RNA polymerase. When an incoming ribonucleoside triphosphate correctly pairs with the next unpaired nucleotide on the DNA template strand, RNA polymerase catalyzes a nucleophilic attack by the 3'-hydroxyl group of the growing RNA strand on the -phosphorus atom of the incoming ribonucleoside triphosphate. As a result, a phosphodiester forms and pyrophosphate is released. The subsequent hydrolysis of pyrophosphate catalyzed by pyrophosphatase provides additional thermodynamic driving force for the reaction. (B and B' represent complementary bases, and hydrogen bonding between bases is indicated by a single dashed line.)
- Figure 21.6 Promoter sequences from 10 bacteriophage and bacterial genes. All these promoter sequences are recognized by the 70 subunit in E. coli. The nucleotide sequences are aligned so that their +1, –10, and –35 regions are in register. Note the degree of sequence variation at each position. The consensus sequence was derived from a much larger database of more than 300 well-characterized promoters.
- Figure 21.13 RNA polymerase II holoenzyme complex bound to a promoter. This model shows various transcription factors bound to RNA polymerase II at a promoter. The transcription factors are often larger and more complex than those shown in this diagram.
- Figure 21.11 A generic eukaryotic promoter showing the basal or “core” promoter elements. The TATA box is described in the text. The BRE is the TFIIB recognition element; while Inr stands for the initiator element. The DPE is the downstream promoter element. The names of the factors that bind to each site are shown above the promoter.
- Figure 21.29 Triose phosphate isomerase gene from maize and the encoded enzyme. (a) Diagram of the gene showing nine exons and eight introns. Some exons contain both translated and untranslated sequences. (b) Three-dimensional structure of the protein showing the parts of the protein encoded by each exon.
- Figure 21.31 Intron removal in mRNA precursors. The spliceosome, a multicomponent RNA-protein complex, catalyzes splicing. [Adapted from Sharp, P. A. (1987). Splicing of messenger RNA precursors. Science 235:766–771.]
- Figure 21.31 Intron removal in mRNA precursors. The spliceosome, a multicomponent RNA-protein complex, catalyzes splicing. [Adapted from Sharp, P. A. (1987). Splicing of messenger RNA precursors. Science 235:766–771.]
- Figure 21.31 Intron removal in mRNA precursors. The spliceosome, a multicomponent RNA-protein complex, catalyzes splicing. [Adapted from Sharp, P. A. (1987). Splicing of messenger RNA precursors. Science 235:766–771.]
- Figure 21.28 Polyadenylation of a eukaryotic mRNA precursor.
- Figure 21.28 Polyadenylation of a eukaryotic mRNA precursor.
- Figure 21.28 Polyadenylation of a eukaryotic mRNA precursor.
- Figure 21.29 Triose phosphate isomerase gene from maize and the encoded enzyme. (a) Diagram of the gene showing nine exons and eight introns. Some exons contain both translated and untranslated sequences. (b) Three-dimensional structure of the protein showing the parts of the protein encoded by each exon.
- Figure 21.21 cAMP production. In the absence of glucose, enzyme III (EIII) transfers a phosphoryl group, originating from phosphoenolpyruvate, to membrane-bound adenylate cyclase. Phosphorylated adenylate cyclase catalyzes the conversion of ATP to cAMP. cAMP binds to CRP, and CRP-cAMP activates the transcription of a number of genes encoding enzymes that compensate for the lack of glucose as a carbon source.
- Figure 21.22 Conformational changes in CRP caused by cAMP binding. Each monomer of the CRP dimer contains a helix-turn-helix motif. In the absence of cAMP, the helices cannot fit into adjacent sections of the major groove of DNA and cannot recognize the CRP-cAMP binding site. When cAMP binds to CRP, the two helices assume the proper conformation for binding to DNA.
- Figure 21.23 Structure of a complex between CRP-cAMP and DNA. Both subunits contain a molecule of cAMP bound at the allosteric site. Each subunit has an helix positioned in the major groove of DNA at the CRP-cAMP binding site. Note that binding induces a slight bend in the DNA. [PDB 1CGP].