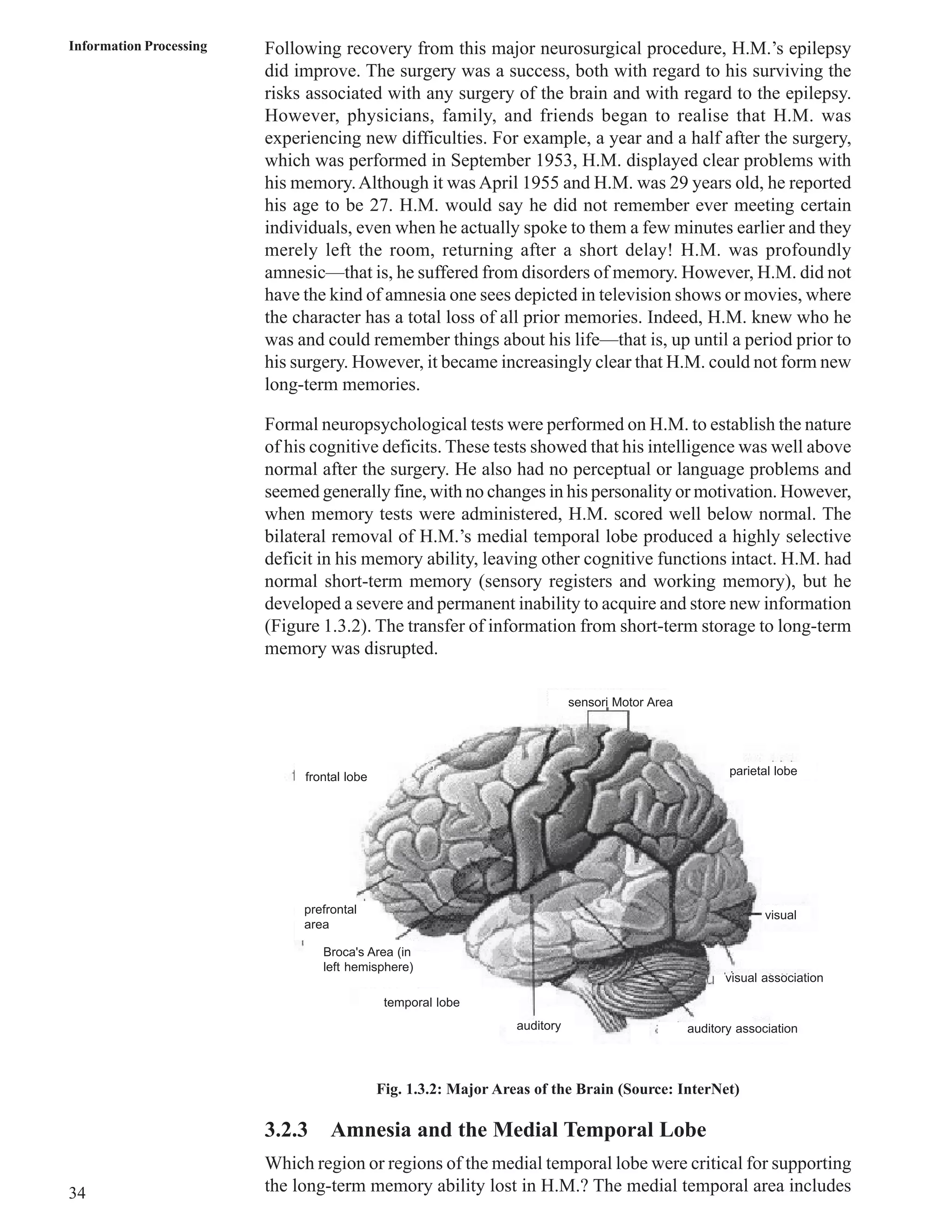
Download to read offline













This document provides an overview of the neuropsychological basis of learning and memory. It discusses how case studies of patients with brain damage or amnesia helped reveal the neural correlates of memory. The case of patient H.M., who had bilateral removal of his medial temporal lobes, showed that this region is critical for forming new long-term memories. Damage limited to the hippocampus, as in patient R.B., also produced severe anterograde amnesia. The hippocampus is involved in consolidating new memories, even though long-term memory storage occurs elsewhere, like the neocortex. Damage to other regions like the diencephalon or anterior temporal lobes can also cause memory impairments by disrupting retrieval or






























![60)NEUROBIOLOGY_OF_MEMORY[1].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/60neurobiologyofmemory1-231214020219-bfbe6909-thumbnail.jpg?width=640&height=640&fit=bounds)




































