Populiacijų genetikayra Mendelio dėsnių, molekulinės
genetikos ir Darvino evoliucijos teorijos sintezė
Dėmesys yra perkeliamas nuo individo į populiaciją, kurios
narys ir yra individas
Kiekvieno geno aleliai populiacijoje sudaro genų bendrą fondą
- pulą (gene pool)
Tik tie individai, kurie reprodukuojasi, dalyvauja kuriant sekančios
kartos genų fondą
Populiacijų genetika tiria genų dažnį bendrame genų fonde ir
kaip jis keičiasi, keičiantis generacijoms
11.1 GENAI POPULIACIJOSE
11-2
3.
Populiacija yravienos rūšies individų grupė, galinti
kryžmintis tarpusavyje
Didelę populiaciją dažniausiai sudaro smulkesnės
grupės, vadinamos subpopuliacijomis
Jos taip pat vadinamos lokaliomis populiacijomis arba
demais
Subpopuliacijos nariams yra didesnė tikimybė
susikryžminti negu bendros populiacijos nariams
Subpopuliacijas dažniausiai skiria nedideli geografiniai
barjerai
Kas yra populiacija?
11-3
4.
Populiacijos yradinaminiai vienetai, kurie keičiasi,
keičiantis generacijoms
Gali pasikeisti populiacijos
1. Dydis
2. Geografinė padėtis
3. Genetinė sudėtis
Populiacijų genetika matematinų teorijų pagalba gali
aprašyti, kaip bendras genų fondas keisis dėl
aukščiau paminėtų fluktuacijų
Kas yra populiacija?
11-4
5.
Terminas polimorfizmasreiškia, kad daugeliui
požymių populiacijoje būdinga variacija
Kai kurie genai yra monomorfiniai, kiti yra
polimorfiniai
11-5
Visi individai priklauso tai pačiai rūšiai Theridion
grallator
Tačiau skiriasi jų aleliai, atsakingi už kūno spalvą ir
raštą
6.
DNR lygmenyjepolimorfizmas atsiranda dėl dviejų
ar daugiau alelių, įtakojančių fenotipą
Polimorfiniais taip pat vadinami genai, kurie
populiacijoje egzistuoja dviejų ir daugiau alelių
forma
Monomorfiniai genai egzistuoja kaip pavieniai
aleliai
Pagal susitarimą, jei tas pats alelis yra randamas bent
99% visų atvejų, genas yra laikomas monomorfiniu
11-6
7.
Alozimai
Allozyme(Alleles of the same gene that encodes an
enzyme)
Tai yra du ar daugiau fermentų (koduojamų to paties
geno), kurių aminorūgščių sekos skiriasi
Tai gali paveikti jų mobilumą elektroforezės metu
Alozimų analizė, naudojant elektroforezę gelyje, ir
DNR sekvenavimas parodė, kad
Daugumoje gamtinių populiacijų didelę genų dalį sudaro
polimorfiniai genai
Tačiau mažose populiacijose, esančiose ant išnykimo
ribos, genetinis kintamumas yra žemas
Taip yra dėl to, kad bendrą genų fondą sudaro nedidelis individų
skaičius
11-7
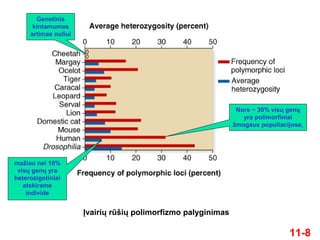
8.
11-8
Įvairių rūšių polimorfizmopalyginimas
Genetinis
kintamumas
artimas nuliui
Nors ~ 30% visų genų
yra polimorfiniai
žmogaus populiacijose,
mažiau nei 10%
visų genų yra
heterozigotiniai
atskirame
individe
9.
Populiacijų genetikojesvarbiausi yra du dydžiai
Populiacijų genetika tiria alelių ir
genotipų dažnius
11-9
Alelių dažnis =
Genotipų dažnis =
Bendras visų šio geno alelių,
esančių populiacijoje, skaičius
Alelių kopijų populiacijoje
skaičius
Bendras visų individų, sudarančių
populiaciją, skaičius
Individų su tam tikru genotipu
skaičius populiacijoje
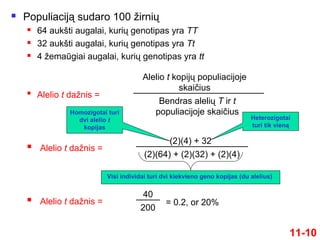
10.
Populiaciją sudaro100 žirnių
64 aukšti augalai, kurių genotipas yra TT
32 aukšti augalai, kurių genotipas yra Tt
4 žemaūgiai augalai, kurių genotipas yra tt
11-10
Alelio t dažnis =
Bendras alelių T ir t
populiacijoje skaičius
Alelio t kopijų populiacijoje
skaičius
Alelio t dažnis =
(2)(64) + (2)(32) + (2)(4)
(2)(4) + 32
Homozigotai turi
dvi alelio t
kopijas
Heterozigotai
turi tik vieną
Visi individai turi dvi kiekvieno geno kopijas (du alelius)
Alelio t dažnis =
200
40
= 0.2, or 20%
11.
Populiaciją sudaro100 žirnių
64 aukšti augalai, kurių genotipas yra TT
32 aukšti augalai, kurių genotipas yra Tt
4 žemaūgiai augalai, kurių genotipas yra tt
11-11
Genotipo tt dažnis tt =
64 + 32 + 4
4
Genotipo tt dažnis =
Bendras individų skaičius
populiacijoje
Genotipo tt augalų skaičius
populiacijoje
Genotipo tt dažnis tt =
100
4
= 0.04, or 4%
% žemaūgių
augalų, esančių
populiacijoje
12.
Kiekvieno požymioalelių ir genotipų dažnis
visą laiką mažesnis arba lygus 1
Monomorfinių genų
Alelio dažnis lygus arba artimas 1.0
Polimorfinių genų
Visų alelių dažnių suma turi būti lygi 1.0
Žirnių populiacijoje
T alelio dažnis + t alelio dažnis = 1
T alelio dažnis = 1 – t alelio dažnis
11-12
= 1 – 0.2
= 0.8, arba 80%
13.
Hardy-Weinbergo lygtįnepriklausomai
vienas nuo kito 1908 m. suformulavo
Godfrey Harold Hardy ir Wilhelm Weinberg
Tai yra paprasta matematinė formulė, susiejanti
alelių ir genotipų dažnius populiacijoje
Hardy-Weinbergo lygtis taip pat dar
vadinama pusiausvyra
Esant tam tikroms sąlygoms alelių ir genotipų
dažnis populiacijoje nesikeičia
11.2 HARDY-WEINBERGO
PUSIAUSVYRA
11-13
14.
Tegul egzistuojadu polimorfinio geno aleliai,
A ir a
Alelio A dažnis yra žymimas kintamuoju p
Alelio a dažnis yra žymimas kintamuoju q
p + q = 1
Šiam genui Hardy-Weinbergo lygtis nustato, kad
(p + q)2
= 1
p2
+ 2pq + q2
= 1
Hardy-Weinbergo lygtis
11-14
Genotipo AA
dažnis
Genotipo Aa
dažnis
Genotipo aa
dažnis
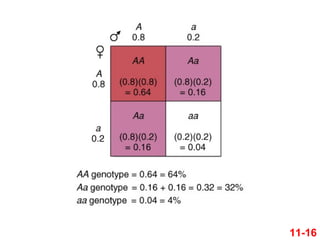
15.
Jei p= 0.8 ir q = 0.2 ir jei populiacija yra Hardy-
Weinbergo pusiausvyroje, tada
AA dažnis = p2
= (0.8)2
= 0.64
Aa dažnis = 2pq
= 2(0.8)(0.2) = 0.32
aa dažnis = q2
= (0.2)2
= 0.04
11-15
Hardy-Weinbergo lygtisnumato pusiausvyrą,
jei populiacija atitinka keletą sąlygų
1. Populiacija yra didelė
Alelių dažniai nesikeičia dėl atsitiktinių imties klaidų
2. Populiacijoje vyksta atsitiktinis kryžminimasis
3. Nėra migracijos
4. Nėra natūraliosios atrankos
5. Nėra mutacijų
Hardy-Weinbergo lygtis nusako kiekybinį ryšį
tarp alelių ir genotipų dažnio
11-17
18.
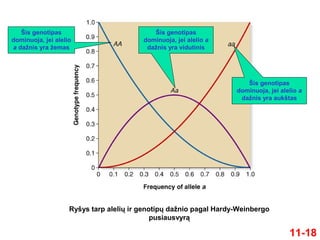
11-18
Ryšys tarp aleliųir genotipų dažnio pagal Hardy-Weinbergo
pusiausvyrą
Šis genotipas
dominuoja, jei alelio
a dažnis yra žemas
Šis genotipas
dominuoja, jei alelio a
dažnis yra aukštas
Šis genotipas
dominuoja, jei alelio a
dažnis yra vidutinis
19.
Realybėje neiviena populiacija pilnai neatitinka
Hardy-Weinbergo pusiausvyros sąlygų
Tačiau kai kurios didelėse gamtinėse populiacijose
migracija yra nedidelė, o natūralioji atranka beveik
nevyksta
Tuo atveju Hardy-Weinbergo pusiausvyros sąlygos kai
kurių genų atžvilgiu yra beveik patenkinamos
Hardy-Weinbergo lygtis gali būti pritaikyta ir 3 ar
daugiau alelių
11-19
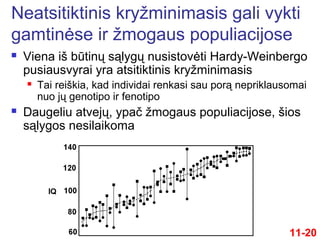
20.
Viena išbūtinų sąlygų nusistovėti Hardy-Weinbergo
pusiausvyrai yra atsitiktinis kryžminimasis
Tai reiškia, kad individai renkasi sau porą nepriklausomai
nuo jų genotipo ir fenotipo
Daugeliu atvejų, ypač žmogaus populiacijose, šios
sąlygos nesilaikoma
Neatsitiktinis kryžminimasis gali vykti
gamtinėse ir žmogaus populiacijose
11-20
21.
Kryžminimasis irfenotipai
Asortatyvus kryžminimasis vyksta tada, kai dažniau
kryžminasi panašių fenotipinių bruožų individai
Disasortatyvus kryžminimasis vyksta tada, kai tarpusavyje
dažniau kryžminasi skirtingų fenotipų individai
Kryžminimasis ir genotipai
Inbrydingas yra genetiškai susijusių individų kryžminimasis
Autbrydingas yra genetiškai nesusijusių individų
kryžminimasis
Nesant kitų evoliucijos veiksnių, inbrydingas ar
autbrydingas nekeičia alelių dažnio
Tačiau dėl tokio kryžminimosi yra pažeidžiamas genotipų balansas
ir jis nebeatitinka Hardy-Weinbergo pusiausvyros
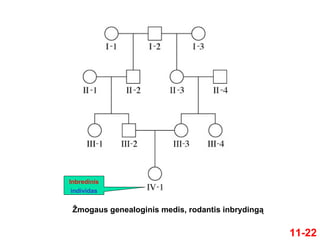
11-21
Vykstant inbrydinguibendras genų fondas yra
mažesnis, nes tėvai yra genetiškai susiję
Wrightas ir Fisheris sukūrė metodus inbrydingo
laipsniui įvertinti
Inbrydingo koeficientas (F) gali būti suskaičiuotas
genealogijoje analizuojant giminingumo laipsnį
11-23
Inbrydingo koeficientas skaičiuojamas taip:
1. Nustatomas bendrų protėvių skaičius
Bendras protėvis yra tas, kuris yra bendras
abiems individo tėvams
IV-1 turi vieną bendrą protėvį I-2
24.
11-24
Jį sudaropenki nariai
2. Nustatomas inbrydingo kelių skaičius
Inbrydingo kelias = trumpiausias genealogijos
kelias, jungiantis abudu tėvus ir bendrą protėvį
Kiekvieno inbrydingo kelio ilgis suskaičiuojamas
sudedant visus individus, sudarančius kelią, išskyrus
tiriamąjį individą
Pateiktame pavyzdyje yra tik vienas kelias
IV-1 III-2 II-2 I-2 II-3 III-3
25.
11-25
3. Skaičiavimamsnaudojama ši formulė
F = Σ (1/2)n
(1 + FA)
Kur
F yra tiriamojo individo inbrydingo koeficientas
n yra individų kiekis inbrydingo kelyje
Išskyrus inbredinį individą
FA yra bendro protėvio inbrydingo koeficientas
Σ rodo sumą dydžių (1/2)n
(1 + FA), paskaičiuotų kiekvienam
inbrydingo keliui
Pateiktame pavyzdyje yra tik vienas bendras protėvis
Kadangi nieko nežinoma apie jo paveldėjimą, daroma prielaida,
kad FA=0
F = Σ (1/2)n
(1 + 0)
= (1/2)5
= 1/32
= 3.125%
Rodo geno homozigotiškumo tikimybę
individe IV-1
Tai atsitinka dėl paveldėjimo iš bendro
protėvio I-2
26.
11-26
Inbrydingo koeficientastaip pat vadinamas
fiksacijos koeficientu
Tai paaiškina simbolį F
Fiksacijos koeficientas yra tikimybė, kad alelis
populiacijoje bus fiksuotas homozigotinėje
būsenoje
27.
11-27
Inbrydingo pasekmėsgali būti įvertintos ir
populiacijoje
Tegul alelių A ir a dažnis yra p ir q
Tada genotipų dažniai bus nusakomi taip
p2
+ Fpq atitinka AA dažnį
2pq(1 – F) atitinka Aa dažnį
q2
+ Fpq atitinka aa dažnį
Tegul p = 0.8, q = 0.2 ir F = 0.25
Tada genotipų dažniai bus
AA = p2
+ Fpq = (0.8)2
+ (0.25)(0.8)(0.2) = 0.68
Aa = 2pq(1 – F) = 2(0.8)(0.2)(1 – 0.25) = 0.24
aa = q2
+ Fpq = (0.2)2
+ (0.25)(0.8)(0.2) = 0.08
28.
11-28
Esant inbrydingui
bus:
68% AA homozigotų
24% heterozigotų
8% aa homozigotų
Nesant inbrydingo
(t.y., F=0) bus:
p2
= 64% AA homozigotų
2pq = 32% heterozigotų
q2
= 4% aa homozigotų
Taigi, inbrydingas didina homozigotų dalį ir mažiną
heterozigotų dalį
Gamtinėse populiacijose inbrydingo koeficientas
didėja, mažėjant populiacijos dydžiui
29.
11-29
Inbrydingas galiturėti tiek teigiamų, tiek ir neigiamų
pasekmių populiacijai
Teigiamos pasekmės stebimos žemės ūkio kultūrose
Inbrydingas gali padidinti proporciją homozigotų, turinčių
reikalingą požymį
Kaip neigiama pasekmė yra tai, kad padidėja
paveldimų recesyvinių ligų
Inbrydingas padidina homozigotiškumo, tuo pačiu ir ligos,
tikimybę
30.
Pagrindinis populiacijųgenetikos klausimas yra
Kaip genetinis polimorfizmas atsiranda ir kaip jis
palaikomas gamtinėse populiacijose?
Labai retiems ir žalingiems aleliams paaiškinimas
yra paprastas
Retos mutacijos sukuria žalingus alelius
Šie žalingi aleliai šalinami iš populiacijos natūraliosios
atrankos būdu
Šie du priešingi procesai palaiko labai žemą žalingų
alelių dažnį
11.3 ALELIŲ DAŽNĮ KEIČIANTYS
VEIKSNIAI
11-30
31.
Tačiau daugeliopolimorfinių alelių dažnis
populiacijoje palaikomas gana aukštas
Šiuo atveju alelių dažniai negali būti paaiškinti naujų
mutacijų atsradimu
Alelių dažniai yra per dideli, o mutacijų – per žemi
Kaip palaikomas aukštas polimorfinių alelių dažnis?
Naujos mutacijos yra alelinės variacijos šaltinis
Po to, kai įvyksta mutacijos, evoliucijos metu šių alelių
dažnis išauga
11-31
32.
Evoliuciniai procesai(arba jėgos) gali būti
suskirstyti į dvi grupes
Neutralūs procesai
Alelių dažnius keičia atsitiktinai
t.y. nepriklausomai nuo individo išgyvenamumo
Migracija ir genų dreifas
Adaptyvūs procesai
Padidina dažnį tų alelių, kurie padeda išgyventi arba
reprodukuotis
Natūralioji atranka
11-32
33.
Mutacijos yrareti atsitiktiniai įvykiai, vykstantys
spontaniškai
Mutagenai padidina mutavimo greitį
Mutacinis kintamumas suteikia žaliavos evoliucijai,
tačiau pats nėra evoliucijos dalis
Populiacijų genetikoje naujas mutacijas
patogiausia nagrinėti būtent tokiame kontekste
Jos paveikia jas paveldinčių individų išgyvenamumą ir
reprodukcinę sėkmę
Mutacijos yra genetinio
kintamumo šaltinis
11-33
34.
Naujos mutacijosgali būti
Naudingos
Neutralios
Žalingos
Neutralių ir žalingų mutacijų atsiradimo tikimybė yra
žymiai didesnė, negu naudingų mutacijų
Mutavimo greitis yra tikimybė, kad gene įvyks
nauja mutacija
Jo dažniausia išraiška yra naujų mutacijų skaičius,
įvykęs tiriamame gene per vieną generaciją
Mutavimo greičio skaitinė išraiška paprastai būna nuo
10-5
iki 10-6
per vieną generaciją
11-34
35.
Kaip mutavimogreitis veikia populiacijos
alelių dažnį?
Tegul geno funkcionalus alelis yra A
Alelio A dažnis žymimas p
Mutacija verčia A į nefunkcionalų alelį a
Alelio a dažnis žymimas q
Mutacijos A a dažnis žymimas u
Reversinės mutacijos (a A) dažnis nykstamai
mažas
11-35
36.
Alelio adažnio padidėjimas per vieną generaciją
yra
∆q = up
Pavyzdžiui,
p = 0.8 (t.y. A dažnis yra 80%)
q = 0.2 (t.y. a dažnis yra 20%)
u = 10–5
(t.y. A mutavimo į a greitis)
∆q = up
= (10–5
)(0.8) = 0.000008
Todėl sekančioje kartoje alelių dažnis bus
qn + 1 = 0.2 + 0.000008 = 0.200008
pn + 1 = 0.8 – 0.000008 = 0.799992
11-36
Taigi, naujos
mutacijos alelių
dažnį vienoje kartoje
keičia labai nežymiai
37.
Alelių dažniopokyčiui suskaičiuoti po daugelio generacijų yra
naudojama formulė
11-37
(1 – u)t
=
pt
p0
Kur
u A mutavimo į a greitis
t yra generacijų skaičius
p0 alelio A dažnis pradinėje generacijoje
pt yra alelio A dažnis po t generacijų
Jei anksčiau nagrinėto pavyzdžio atveju norime sužinoti,
koks bus alelio dažnis po 1000 generacijų, tada
(1 – 0.00001)1000
=
pt
0.8
pt = 0.792
Taigi, po 1000 generacijų alelio A
dažnis sumažės nuo 0.8 tik iki 0.792
Tai vėl rodo, kaip lėtai mutacijos
keičia alelių dažnį
38.
Atsitiktinis genųdreifas yra atsitiktinis alelių dažnio
pokytis, atsirandantis dėl riboto gametų
pasirinkimo
Kitaip tariant alelių dažnis gali keistis iš kartos į
kartą dėl atsitiktinių priežasčių
Dėl genų dreifo aleliai gali būti arba fiksuoti
populiacijoje, arba pamesti iš viso
Genų dreifo įtaka alelių dažniui priklauso nuo
populiacijos dydžio
Genų dreifas
11-38
39.
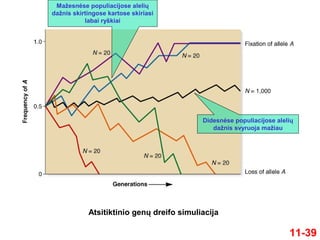
11-39
Atsitiktinio genų dreifosimuliacija
Mažesnėse populiacijose alelių
dažnis skirtingose kartose skiriasi
labai ryškiai
Didesnėse populiacijose alelių
dažnis svyruoja mažiau
40.
Jei individasturi dvi tiriamojo geno kopijas, tada
tikėtinas naujų mutacijų skaičius = 2Nu,
kur u yra mutavimo greitis
N yra individų skaičius populiacijoje
Tai reiškia, kad naujos mutacijos greičiau atsiras
didelėse populiacijose, lyginant su mažomis
11-40
Kiek naujų mutacijų galima tikėtis gamtinėse
populiacijose?
41.
Tikimybė užfiksuotinaują mutaciją dėl genų dreifo yra:
fiksacijos tikimybė = 1/2N
Daroma prielaida, kad abiejų lyčių indėlis į sekančią generaciją yra
vienodas
Kitaip tariant, fiksacijos tikimybė yra tokia pati, kaip ir alelių
dažnis populiacijoje
Pavyzdžiui, jei N = 20, tada
fiksacijos tikimybė = 1/(2 X 20), or 2.5%
Priešingai, naujas alelis gali būti išeliminuotas iš
populiacijos
Eliminacijos tikimybė = 1 – 1/2N
11-41
Kokia tikimybė, kad nauja mutacija bus užfiksuota
arba išeliminuota iš populiacijos dėl genų dreifo?
42.
Populiacijos dydisN turi priešingą poveikį
mutacijoms ir jų fiksacijai
Kai N yra labai didelis
Naujos mutacijos labiau tikėtinos
Tačiau kiekviena nauja mutacija turi didesnę tikimybę
būti eliminuota iš populiacijos dėl genų dreifo
Kai N yra labai mažas
Naujos mutacijos populiacijoe atsiranda labai retai
Tačiau kiekvienai nauja mutacija turi didesnę tikimybę
būti fiksuota populiacijoje dėl genų dreifo
11-42
43.
Tai vėlpriklauso nuo individų skaičiaus populiacijoje:
11-43
t = 4N
kur
t yra vidutinis generacijų skaičius, reikalingas fiksacijai pasiekti
N yra individų skaičius populiacijoje, darant prielaidą, kad abiejų
lyčių indėlis į sekančią generaciją yra vienodas
Kaip ir galima tikėtis, alelių fiksacijai reikės žymiai
daugiau laiko didelėse populiacijose
Jei populiacijoje yra 1 milijonas besikryžminančių individų, reikės
4 milijonų generacijų fiksacijai įvykti
Jei populiacijoje bus tik 100 individų, fiksacijai reikės 400
generacijų
Kiek generacijų reikia mutacijai užfiksuoti?
44.
Genų dreifaspasižymi dviem svarbiomis
ypatybėmis
1. Genų dreifas tam tikro alelio dažnio požiūriu
veikia kryptingai
Galiausiai alelis arba fiksuojamas populiacijoje, arba
išeliminuojamas
2. Genų dreifo įtaka didesnė mažose
populiacijose
Genų dreifo pavyzdžiai
Butelio kaklelio efektas
Įkūrėjo (pradininko) efektas
11-44
45.
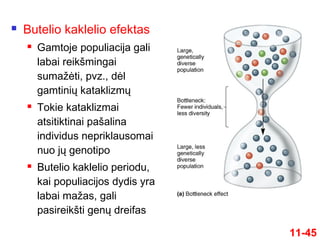
11-45
Butelio kaklelioefektas
Gamtoje populiacija gali
labai reikšmingai
sumažėti, pvz., dėl
gamtinių kataklizmų
Tokie kataklizmai
atsitiktinai pašalina
individus nepriklausomai
nuo jų genotipo
Butelio kaklelio periodu,
kai populiacijos dydis yra
labai mažas, gali
pasireikšti genų dreifas
46.
Afrikinis gepardasprarado beveik visą savo genetinį kintamumą
Taip atsitiko dėl butelio kaklelio efekto, įvykusio prieš 10,000 - 12,000 metų
11-46
47.
11-47
Įkūrėjo efektas
Nedidelė individų grupė atsiskiria nuo didesnės
populiacijos ir įkuria koloniją naujoje vietoje
Tai turi dvi svarbias pasekmes
1. Tikėtina, kad įkurtoji populiacija pasižymės mažesniu
genetiniu kintamumu, negu pirminė populiacija
2. Įkurtosios populiacijos alelių dažniai gali žymiai skirtis nuo
pirminės populiacijos
Pavyzdys – Lankasteryje (JAV) gyvena menonitų
sektos bendruomenė, kuriai būdinga socialinė izoliacija
– bendruomenės nariai šeimas gali kurti tik tarpusavyje
Ją sudaro apie 8000 asmenų, kilusių iš trijų porų, kurios 1770
m. įmigravo į JAV
Ellis-van Creveld sindromo (liliputiškumo recesyvinė forma)
dažnis yra 7%
Tai daug daugiau nei bet kurioje kitoje populiacijoje
48.
Migracija tarpdviejų skirtingų populiacijų gali
pakeisti alelių dažnį
Naujoji populiacija (recipientinė populiacija) yra
vadinama konglomeratu
Norint suskaičiuoti alelių dažnius konglomerate reikia
žinoti
1. Pradinį alelių dažnį donorinėje ir recipientinėje populiacijose
2. Konglomeratinės populiacijos dalį, kurią sudaro migrantai
Migracija
11-48
49.
∆pC =m(pD – pR)
11-49
Kur
∆pC alelio dažnio pokytis konglomeratinėje populiacijoje
pD alelio dažnis donorinėje populiacijoje
pR is alelio dažnis pradinėje recipientinėje populiacijoje
m migrantų dalis konglomeratinėje populiacijoje
m =
Donorų skaičius
konglomeratinėje populiacijoje
Bendras individų skaičius
konglomeratinėje populiacijoje
50.
∆pC =m(pD – pR)
= 0.2(0.7 – 0.3)
11-50
Pavyzdys:
Alelio A dažnis yra 0.7 donorinėje ir 0.3 recipientinėje
populiacijose
20 individų migruoja ir prisijungia prie recipientinės
populiacijos, kurią sudaro 80 individų
m =
20
20 + 80
= 0.2
= 0.08
pC = pR + ∆pC =
0.3 + 0.08
= 0.38
Alelio A dažnis išaugo
nuo 0.3 iki 0.38
51.
11-51
Populiacijų genetikąlabiau domina genų srautas,
t.y. ne tiek individų migracija, kiek alelių dažnio pokyčiai
Gamtoje individai tarp populiacijų dažniausiai
migruoja abiem kryptimis
Ši dvikryptė migracija turi svarbias pasekmes
1. Ji mažina alelių dažnio skirtumus tarp populiacijų
2. Ji skatina genetinę įvairovę populiacijos viduje
52.
Charles Darwinir Alfred Russel Wallace
nepriklausomai vienas nuo kito sukūrė
natūraliosios atrankos teoriją
Pagal šią teoriją egzistuoja kova dėl išgyvenimo
Tie individai, kurie yra geriau prisitaikę prie tam tikros
aplinkos, išgyvena ir reprodukuojasi
Dabar aišku, kad natūralioji atranka taip pat
susijusi su dauginimosi efektyvumu ir fertilumu, o
ne tik su skirtingu išgyvenamumu
Natūralioji atranka palankesnė
labiau prisitaikiusiems
11-52
53.
Natūraliosios atrankosšiuolaikinis aprašymas sieja
molekulinę genetiką su individų fenotipais
1. Populiacijos viduje yra genetinė variacija, atsirandanti
dėl DNR sekų skirtumų
Skirtingi aleliai gali koduoti besiskiriančių funkcijų baltymus
2. Kai kurie aleliai gali koduoti baltymus, padidinančius
individų išgyvenamumą ar reprodukcines galimybes
3. Individų, turinčių naudingus alelius, tikimybė išgyventi ir
reprodukuotis yra didesnė
4. Per daugelį generacijų daugelio skirtingų genų alelių
dažnis gali pasikeisti dėl natūraliosios atrankos
Tai reikšmingai keičia rūšies savybes
Galutinis natūraliosios atrankos rezultatas yra populiacija, geriau
prisitaikiusi prie aplinkos ir/ar efektyviau besidauginanti
11-53
54.
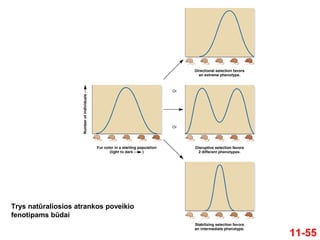
Natūralioji atrankaveikia fenotipus (kurie priklauso
nuo individo genotipo)
Natūralioji atranka kiekybinius požymius veikia
trimis būdais
1. Kryptinga atranka
Padeda išgyventi vienam kraštutiniam fenotipui, kuris yra
geriausiai prisitaikęs prie aplinkos sąlygų
2. Disruptyvioji (arba diversifikuojanti) atranka
Padeda išgyventi dviem (ar daugiau) skirtingų fenotipų
3. Stabilizuojanti atranka
Padeda išgyventi individams su tarpiniu fenotipu
11-54
Natūraliosios atrankoskiekybinių pasekmių
aptarimą reikia pradėti nuo Darvininio
prisitaikomumo (Darwinian fitness) sąvokos
Prisitaikomumas yra santykinė tikimybė to, kad individas
išgyvens ir įneš indėlį į sekančios kartos bendrą genų
fondą
Darvininis prisitaikomumas yra reprodukcinio
pranašumo matas
Tegul genas turi du alelius, A ir a
Trims genotipų klasėms gali būti priskirtos
prisitaikomumo reikšmės, nustatomos pagal jų
reprodukcinį potencialą
11-56
57.
Tegul vidutinėreprodukcinė sėkmė yra:
AA 5 palikuonys
Aa 4 palikuonys
aa 1 palikuonis
Genotipui, pasižyminčiam didžiausia reprodukcine
sėkme, priskiriama prisitaikomumo reikšmė, lygi
1.0
Kitų genotipų prisitaikomumas nustatomas santykinai,
geriausiai prisitaikiusio genotipo atžvilgiu
Prisitaikomumo reikšmės žymimos simboliu W
Genotipo AA prisitaikomumas: WAA = 1.0
Genotipo Aa prisitaikomumas: WAa = 4/5 = 0.8
Genotipo aa prisitaikomumas: Waa = 1/5 = 0.2
11-57
58.
Reprodukcinės sėkmėsskirtumai gali būti dėl to, kad
1. Labiausiai prisitaikę fenotipai geriau išgyvena
2. Labiausiai prisitaikę fenotipai dažniau susiranda
reprodukcinius partnerius
3. Labiausiai prisitaikę fenotipai yra fertilesni
Priešingas prisitaikomumui parametras yra atrankos
(selekcijos) koeficientas (s)
Jis matuoja, kaip efektyviai genotipas yra šalinamas iš
populiacijos
s = 1 – W
11-58
59.
Kaip prisitaikomumasveikia Hardy-Weinbergo
pusiausvyrą ir alelių dažnius?
Egzistuoja du geno aleliai, A ir a
Jų prisitaikomumo reikšmės yra
WAA = 1.0
WAa = 0.8
Waa = 0.2
Sekančioje kartoje Hardy-Weinbergo pusiausvyra bus
modifikuota taip:
Genotipo AA dažnis: p2
WAA
Genotipo Aa dažnis: 2pqWAa
Genotipo aa dažnis: q2
Waa
11-59
60.
Šių trijųkomponentų suma nebūtinai turi būti lygi 1.0,
kaip kad esant Hardy-Weinbergo pusiausvyrai
Vietoje to, jų suma yra lygi dydžiui, vadinamam vidutiniu
populiacijos prisitaikomumu
11-60
p2
WAA + 2pqWAa + q2
Waa = W
Abi lygties puses galima padalinti iš vidutinio
populiacijos prisitaikomumo
p2
WAA
W
2pqWAa
W
+
q2
Waa
W
+ = 1
Dabar galima apskaičiuoti tikėtinus genotipų ir alelių
dažnius po vienos generacijos, veikiant natūraliajai atrankai
61.
AA genotipodažnis =
11-61
p2
WAA
W
Aa genotipo dažnis =
2pqWAa
W
aa genotipo dažnis =
q2
Waa
W
Alelio A dažnis: pA =
p2
WAA
W
pqWAa
W
+
Alelio a dažnis: qa =
q2
Waa
W
pqWAa
W
+
62.
Pavyzdys:
Pradinisalelių dažnis yra A = 0.5 ir a = 0.5
Prisitaikomumo reikšmės yra 1.0, 0.8 ir 0.2 genotipams
AA, Aa ir aa
11-62
p2
WAA + 2pqWAa + q2
Waa = W
Po vienos kartos, veikiant natūraliajai atrankai,
turėsime:
W = (0.5)2
(1) + 2(0.5)(0.5)(0.8) + (0.5)2
(0.2)
= 0.25 + 0.4 + 0.05
= 0.7
63.
AA genotipodažnis =
11-63
p2
WAA
W
Aa genotipo dažnis =
2pqWAa
W
aa genotipo dažnis =
q2
Waa
W
Alelio A dažnis: pA =
p2
WAA
W
pqWAa
W
+
Alelio a dažnis: qa =
q2
Waa
W
pqWAa
W
+
=
(0.5)2
(1)
0.7
= 0.36
= 0.57
2(0.5)(0.5)(0.8)
0.7
=
=
(0.5)2
(0.2)
0.7
= 0.07
= 0.36 + 0.57/2 = 0.64
= 0.07 + 0.57/2 = 0.36
64.
Sekančioje kartoje
f(A) išauga nuo 0.5 iki 0.64
f(a) sumažėja nuo 0.5 iki 0.36
Taip yra dėl to, kad AA genotipui būdingas didžiausias
prisitaikomumas
Kitas įdomus natūraliosios atrankos bruožas yra tas, kad ji
didina vidutinį populiacijos prisitaikomumą
Padarę prielaidą, kad individualios prisitaikomumo reikšmės yra
pastovios, gauname:
11-64
p2
WAA + 2pqWAa + q2
WaaW =
= (0.64)2
(1) + 2(0.64)(0.36)(0.8) + (0.36)2
(0.2)
= 0.80
Vidutinis populiacijos prisitaikomumas
išaugo nuo 0.7 iki 0.8
65.
Pasinaudojus tapačia procedūra, galima
apskaičiuoti reikšmes, stebimas dar kitoje
kartoje
f(A) išaugs iki 0.85
f(a) sumažės iki 0.15
Vidutinis populiacijos prisitaikomumas išaugs iki
0.931
Taigi, bendra tendencija yra tokia, kad A dažnis
auga, a dažnis mažėja, vidutinis populiacijos
prisitaikomumas taip pat auga
11-65
66.
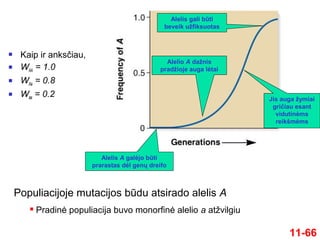
11-66
Kaip iranksčiau,
WAA = 1.0
WAa = 0.8
Waa = 0.2
Alelis A galėjo būti
prarastas dėl genų dreifo
Alelio A dažnis
pradžioje auga lėtai
Jis auga žymiai
gričiau esant
vidutinėms
reikšmėms
Alelis gali būti
beveik užfiksuotas
Populiacijoje mutacijos būdu atsirado alelis A
Pradinė populiacija buvo monorfinė alelio a atžvilgiu
67.
Polimorfizmas galipasiekti pusiausvyrą, jei
priešingos atrankos jėgos atsveria viena kitą
Populiacija nesivysto alelio eliminacijos ar fiksacijos
kryptimis
Tokia situacija vadinama subalansuotu polimorfizmu
Jis gali atsirasti dėl skirtingų priežasčių
1. Selektyvaus heterozigotų pranašumo
2. Rūšies užimamo arealo, sudaryto iš skirtingų aplinkų
Subalansuotas polimorfizmas
11-67
68.
1. Heterozigotaituri selektyvų pranašumą
Didesnis heterozigotų prisitaikomumas yra
balansuojamas žemesnio abiejų homozigotų
prisitaikomumo
Pavyzdys:
Prisitaikomumas:
WAA = 0.7
WAa = 1.0
Waa = 0.4
11-68
Atrankos koeficientai:
sAA = 1 – 0.7 = 0.3
sAa = 1 – 1.0 = 0
saa = 1 – 0.4 = 0.6
Tai yra
heterozigotų
pranašumo
pavyzdys
Alelio A dažnis =
Saa
sAA + saa
0.6
0.3 + 0.6
= = 0.67
Esant tokioms sąlygoms, populiacija pasieks
pusiausvyrą, kurioje:
Alelio a dažnis =
SAA
sAA + saa
= 0.33
0.3
0.3 + 0.6
=
69.
Subalansuotas polimorfizmaskartais gali
paaiškinti aukštą dažnį alelių, kurie yra
kenksmingi homozigotinėje būsenoje
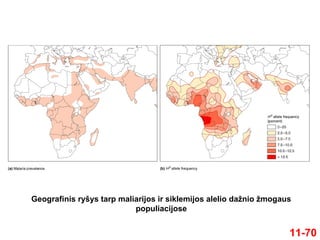
Pavyzdys: žmogaus β-globino geno HbS
alelis
HbS
HbS
sukelia siklemiją
HbA
HbA
yra fenotipiškai normalūs
HbA
HbS
pasižymi geriausiu prisitaikomumu endeminiuose
maliarijos regionuose
Heterozigotų šansai išgyventi, užsikrėtus parazitu Plasmodium
falciparum, yra geresni
Kiti heterozigotų pranašumo pavyzdžiai yra
Cistinė fibrozė
Heterozigotai atsparūs diarėjinėms ligoms (pvz., cholerai)
Tay-Sachso liga
Heterozigotai atsparesni tuberkuliozei
11-69
11-71
2. Rūšisužima arealą, kurį sudaro skirtingos
gamtinės ir ekologinės sritys
Vienomis sąlygomis pranašesnis gali būti vienas
alelis, kitomis - kitas
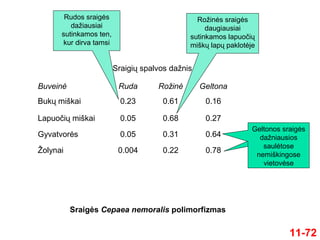
Sraigė Cepaea nemoralis
gyvena tiek miškuose, tiek ir
atvirose vietovėse
Šiai sraigei būdingas spalvos ir
kriauklės rašto polimorfizmas
1954 m. A. J. Cain ir P. M.
Sheppard nustatė, kad sraigės
spalva koreliuoja su jos
gyvenamąja aplinka
72.
11-72
Buveinė Ruda RožinėGeltona
Bukų miškai 0.23 0.61 0.16
Lapuočių miškai 0.05 0.68 0.27
Gyvatvorės 0.05 0.31 0.64
Žolynai 0.004 0.22 0.78
Sraigės Cepaea nemoralis polimorfizmas
Sraigių spalvos dažnis
Rudos sraigės
dažiausiai
sutinkamos ten,
kur dirva tamsi
Rožinės sraigės
daugiausiai
sutinkamos lapuočių
miškų lapų paklotėje
Geltonos sraigės
dažniausios
saulėtose
nemiškingose
vietovėse
73.
Genetinio kintamumopozityvioji pusė yra ta,
kad jis suteikia įdomumo ir žavesio mūsų
gyvajai gamtai
Negatyvioji pusė yra ta, kad kai kurie
genetinio kintamumo tipai yra žalingi
populiacijos išgyvenamumui
Genetinis krūvis yra genetinio
kintamumo neigiama pasekmė
11-73
74.
Genetinis krūvis(L; genetic load) yra genetinis
kintamumas, mažinantis vidutinį populiacijos
prisitaikomumą, lyginant su (teorine) maksimalia ar
optimalia reikšme
Genetinis krūvis yra genetinio
kintamumo neigiama pasekmė
11-74
L =
Wmax
W(Wmax – )
kur
L yra genetinis krūvis
W
yra vidutinis populiacijos prisitaikomumas
Wmax yra maksimalus populiacijos prisitaikomumas
75.
Daugelis veiksniųkuria populiacijos genetinį krūvį
Tai yra:
1. Mutacijos
2. Segregacija
3. Rekombinacija
4. Aplinkos heterogeniškumas
5. Mejozinis draivas/gametų aranka
6. Motinos-vaisiaus nesuderinamumas
7. Ribotas populiacijos dydis
8. Migracija
11-75